
Les plantes font partie de la lignée végétale, c’est-à-dire d’un ensemble d’organismes autotrophes et phototrophes .
La lignée végétale traverse divers règnes. Elle prend naissance au sein des procaryotes avec les cyanobactéries (algues bleues) et les chloroxybactéries. Elle inclut de nombreux organismes appartenant au règne des protistes/protoctistes (rassemblés communément sous le vocable « algues« ) et s’épanouit avec les plantes « supérieures » (des Bryophytes aux Angiospermes). Selon ces classifications, les plantes sont également appelées Embryophytes, Cormophytes ou Archégoniates.
Définir une plante, de manière simple, est chose plus malaisée qu’il n’y paraît. S’il est vrai qu’en principe les plantes ont en commun un certain nombre de caractères, elles ne les possèdent pas nécessairement de manière exclusive et par ailleurs ne les possèdent pas nécessairement tous. (Photo 1). Considérées individuellement, ces caractéristiques ne constituent pas toujours un élément décisif de classement. Nous considérerons successivement:
- Le cormus
- Les sporanges et les gamétanges
- L’oogamie
- L’hétérogamie
- L’embryon
- Le cycle de développement digénétique haplodiplophasique
- Les pigments et les substances de réserve
- Les adaptations à la vie terrestre
 Photo 1 : Cuscuta europaea, une angiosperme sans racine.
Photo 1 : Cuscuta europaea, une angiosperme sans racine.
1. Le cormus
Au sein de la lignée végétale, une distinction est opérée entre Thallophytes et Cormophytes (cf. Figure 1).
Les premiers possèdent un appareil végétatif, le thalle, dépourvu de tiges feuillées caractérisées, même au maximum de sa complexité. L’appareil végétatif des seconds, le cormus, est nécessairement constitué de rameaux feuillés et souvent de racines, édifiés par des méristèmes apicaux.

Figure 1- cladogramme des Cormophytes (ou Embryophytes)
Sur la base de ce critère organographique, les procaryotes et les protistes/protoctistes sont tous des thallophytes (de même que les mycètes pour ceux qui persisteraient à les classer dans la lignée végétale) bien que des convergences morphologiques soient susceptibles de donner au thalle, un aspect qui ressemble à une feuille ou à un rameau feuillé. C’est le cas notamment d’algues brunes (Photo 1), rouges ou vertes.
 Photo 1 : Fucus vesiculosus, une algue brune (= Phaeophyta).
Photo 1 : Fucus vesiculosus, une algue brune (= Phaeophyta).
Par ailleurs, dans les Embryophytes, les Bryophytes (au sens large, comprenant les Anthocerophyta, les Marchantiophyta et les Bryophyta), classé parmi les Cormophytes, se situe en fait au niveau de la transition Thallophytes et Cormophytes. En effet, chez les Anthoceros et diverses hépatiques, le cormus a subi une régression qui l’a rendu thalloïde (on parle de cormus thalloïde ou même, fucoïde (ressemblant à un fucus) (Photo 2) alors que, dans la classe des Bryophytes (les mousses), le gamétophyte a l’aspect d’un cormus ramifié (tige = caulidie) et feuillé (feuille = phyllidie). Le sporophyte, quant à lui, se présente sous la forme d’un axe rudimentaire, sans ramification ni vascularisation.
Les Trachéophytes comprennent les Ptéridophytes et les Spermatophytes (Photo 3).
Pour la clarté de l’exposé qui suit, nous désignerons le règne des plantes, Bryophytes compris, par le vocable « Cormophytes » et l’ensemble des groupes considérés comme plus primitifs constituera les « Thallophytes« .
 Photo 2 : Marchantia polymorpha, une hépatique à thalle
Photo 2 : Marchantia polymorpha, une hépatique à thalle
 Photo 3 : Crassula tiliae, la plus petite plante à fleur terrestre
Photo 3 : Crassula tiliae, la plus petite plante à fleur terrestre
2. Les sporanges et les gamétanges
La structure des organes reproducteurs fournit la possibilité de distinguer thallophytes et cormophytes.
a. Les Thallophytes
Au moment de la reproduction, les thallophytes eucaryotes produisent des spores et des gamètes.
La spore (du grec ancien σπορά, « ensemencement, semence ») est une cellule spécialisée, capable de donner naissance à un nouvel organisme par divisions mitotiques, sans qu’il y ait fusion préalable avec une autre cellule. L’organisme, issu de la spore, est le résultat d’une reproduction agame (c’est-à-dire n’impliquant pas la fusion de gamètes) et peut donc se perpétuer, identique à lui-même, de générations en générations. La spore assure aussi bien la propagation des formes haploïdes, lorqu’elle est le produit de la méiose, que des formes diploïdes lorsqu’elle est produite en dehors de tout phénomène de méiose (reproduction asexuée).
Le gamète est une cellule reproductrice, mâle ou femelle (ou de signe + ou -) dont le noyau ne contient qu’un seul chromosome de chaque paire (haploïde) et qui ne peut se multiplier seule, mais doit s’unir à un gamète de sexe opposé (de signe opposé), dans l’acte de fécondation, pour donner un zygote qui se développera en un nouvel organisme, résultat de la reproduction sexuée. Le zygote et le nouvel organisme auquel il donne naissance possèdent donc un double jeu de chromosomes (diploïde).
Pour qu’un gamète puisse à nouveau se former, un retour à l’état haploïde est requis. La réduction précise du nombre de chromosomes, réalisant le partage exact du matériel génétique, s’effectue au cours de la méiose. Celle-ci est donc liée à l’installation même de la reproduction sexuée et de la fécondation qui la caractérise, car elle est le mécanisme compensateur de cette dernière. L’alternance méiose-fécondation introduit donc un cycle dans le développement d’un organisme puisqu’on peut distinguer une période où les noyaux des cellules sont haploïdes et une période où ils sont diploïdes. L’importance relative de ces deux périodes n’est pas la même chez tous les organismes, méiose et fécondation pouvant être plus ou moins séparées et situées en des points différents du cycle de développement.
Ces spores et ces gamètes sont logés respectivement dans des sporocystes et des gamétocystes issus d’une cellule-mère (Figure 1). Le noyau de celle-ci subit plusieurs divisions successives et chaque noyau fils sera celui d’une spore ou d’un gamète.
Chez les thallophytes, le sporocyste ou gamétocyste, renfermant les spores ou les gamètes, est donc limité par la paroi de la cellule-mère.
b. Les Cormophytes
Chez les cormophytes, au cours des premières divisions d’une cellule-mère, une (ou plusieurs) assise(s) pluricellulaire(s) limitant l’organe au sein duquel se produiront spores ou gamètes est (sont) mise(s) en place (Figure 1). Les sporanges et les gamétanges sont donc limités par une ou plusieurs couches de cellules.
 Figure 1 : Structure des organes reproducteurs des thallophytes et des cormophytes (y compris les Bryophytes)
Figure 1 : Structure des organes reproducteurs des thallophytes et des cormophytes (y compris les Bryophytes)
Chez les Cormophytes (y compris les Bryophytes), la sporogenèse se déroule habituellement selon un même schéma (Figure 2) avec quelques variantes qui seront, le cas échéant, signalées. Une (ou plusieurs) cellule(s) du sporange, appelée(s) l'(les) archéspore (s), est (sont) à l’origine des cellules reproductrices agames.
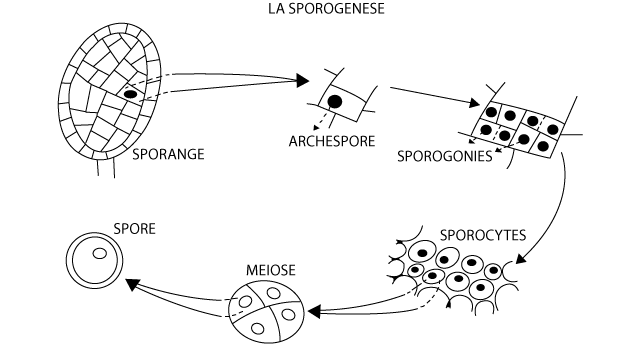 Figure 2 : La sporogenèse chez les plantes
Figure 2 : La sporogenèse chez les plantes
Chaque archéspore se divise et produit un certain nombre de cellules-mères de spores ou sporogonies ; chaque sporogonie grandit et s’individualise en se séparant de ses voisines pour évoluer en sporocyte.
Les sporocytes subissent la méiose et donnent quatre cellules à noyau réduit qui restent tout d’abord associées en tétrade. Chaque élément de la tétrade s’isole ensuite (parfois restent accolé (chez certaines hépatiques)) pour former une spore (méiospore).
Les gamétanges mâles (produisant les gamètes mâles) et femelles (renfermant chacun un gamète femelle ou oosphère) sont appelés respectivement anthéridies et archégones.
4. L’hétérogamie
Chez les végétaux, la forme des gamètes varie beaucoup. Dans un même groupe de plantes à thalle (soit les algues brunes, soit les algues vertes, etc.), on trouve des formes à gamètes mâles et femelles mobiles (isogamie ou anisogamie) (Figure ?), soit des formes où le gamète femelle est immobile (oogamie). Enfin, de rares formes où le gamète mâle devient à son tour immobile (oogamie siphonogame de quelques ex-champignons, comme les Oomycètes (aujourd’hui classés dans les algues) (Figure 3).
Les Cormophytes produisent toujours des gamètes mâles et femelles morphologiquement différents. Il y a donc nécessairement hétérogamie. Celle-ci paraît inévitable puisque le gamète femelle reste fixé sur l’organisme qui le produit : il faut nécessairement que les gamètes mâles soient libérés par les organes qui les produisent et soient mobiles pour que la fécondation soit possible. L’hétérogamie apparaît déjà au sein des algues. Un des cas les plus démonstratifs est celui des Charophytes (Figure 4).
 Figure 3 – oogamie siphonogame chez le mildiou de la vigne (Oomycètes)
Figure 3 – oogamie siphonogame chez le mildiou de la vigne (Oomycètes)
 Figure 4: Cycle hétéromorphe (3 générations dissemblables) et hétérogamique (gamètes mâle = spermatie – femelle = carpogonium) d’une algue rouge
Figure 4: Cycle hétéromorphe (3 générations dissemblables) et hétérogamique (gamètes mâle = spermatie – femelle = carpogonium) d’une algue rouge
6. Le cycle de développement digénétique haplodiplophasique
Les plantes possèdent un cycle de développement typiquement digénétique haplodiplophasique, c’estéàédire où alternent toujours deux générations distinctes (une génération étant représentée par l’existence d’un individu) :
Chaque nouvelle génération est issue d’une cellule isolée : la méiose qui donne le gamétophyte ou le zygote qui se développe en sporophyte. L’importance de chacune de ces générations et leurs rapports varient considérablement selon les groupes (embranchements) d’organismes considérés.
La reproduction des algues: Des cycles de développement variés et parfois inconnus chez les plantes ont été expérimentés par les algues et sans doute est-il utile de rappeler ici ces différentes expériences afin de permettre une meilleure compréhension du cycle de reproduction des plantes.
Chez les algues, la reproduction agame, c’est-à-dire ne mettant pas en jeu la fusion de gamètes dans l’acte de fécondation, reste bien présente.
La reproduction sexuée, absente ou rare dans les groupes les plus primitifs, s’impose définitivement parmi les groupes les plus évolués. Les gamètes mâles et femelles sont soit identiques (+ et -) et l’on parle alors d’isogamie ; soit de même forme mais de taille différente (anisogamie), soit de forme différente (hétérogamie).
Les cycles de reproduction sont monogénétiques haplophasiques (Figure 5) lorsque la méiose est corrective, c’est-à-dire lorsqu’elle survient immédiatement après la fécondation et que le cycle de développement est caractérisé par la présence d’un seul organisme haploïde, issu de la méiospore.
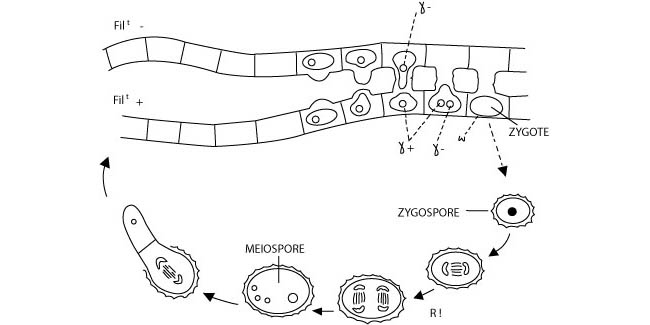 Figure 5 : Le cycle de reproduction monogénétique haplophasique chez Spyrogyra, une algue verte
Figure 5 : Le cycle de reproduction monogénétique haplophasique chez Spyrogyra, une algue verte
Ils sont monogénétiques diplophasiques (Figure 6) lorsque la méiose est préventive, c’est-à-dire lorsqu’elle est suivie immédiatement de la fécondation et que le cycle de développement est caractérisé par la présence d’un seul organisme diploïde, issu du zygote.
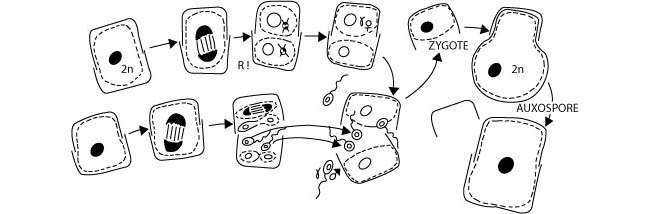 Figure 6 : Le cycle de reproduction monogénétique diplophasique d’une diatomée, une algue unicellulaire
Figure 6 : Le cycle de reproduction monogénétique diplophasique d’une diatomée, une algue unicellulaire
Ils sont digénétiques haplodiplophasiques lorsque méiose et fécondation sont séparées par des phases de divisions mitotiques donnant chacune naissance à un organisme résultant donc, l’un d’une reproduction agame, l’autre de la reproduction sexuée (Figures 7 et 8).
N.B: Les Rhodophytes (algues rouges) (Photo 4) présentent des cycles trigénétiques avec deux types de sporophytes dont un seul réalisera la méiose, l’autre formant des spores végétatives 2N.
 Photo 4 : Une algue rouge encroûtante ; Ralfsia verrucosa.
Photo 4 : Une algue rouge encroûtante ; Ralfsia verrucosa.
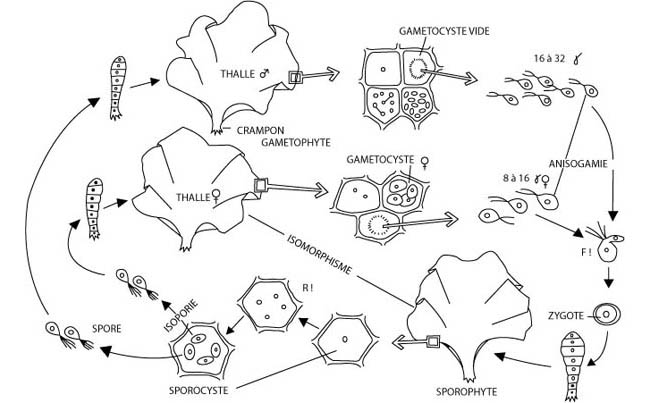 Figure 7 : Le cycle de reproduction digénétique haplodiplophasique de Ulva lactuca, une algue verte
Figure 7 : Le cycle de reproduction digénétique haplodiplophasique de Ulva lactuca, une algue verte
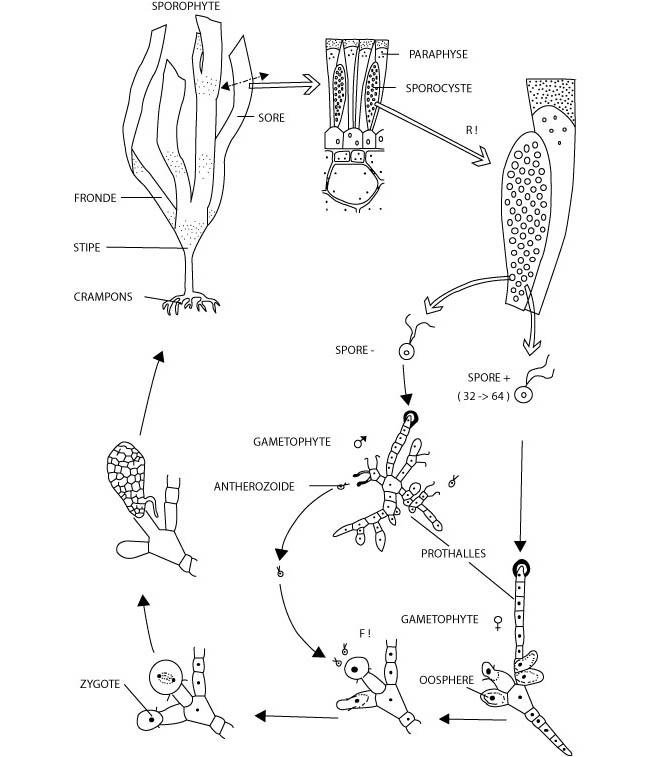 Figure 8: Le cycle de reproduction digénétique haplodiplophasique de Laminaria sp., une algue brune
Figure 8: Le cycle de reproduction digénétique haplodiplophasique de Laminaria sp., une algue brune
7. Les pigments et les substances de réserves
Tous les Embryophytes (ainsi que les algues vertes) possèdent des chloroplastes (Figure 9) contenant, outre des chlorophylles a comme tous les Eucaryotes photosynthétiques, les cyanobactéries et quelques bactéries sulfureuses vertes, des chlorophylles b (Figure 9) et différents autres pigments, dont les carotènes présents chez tous les organismes photosynthétiques. Les substances de réserve sont généralement constituées par de l’amidon (1-4 glucane) intraplastidial.
Les autres organismes de la lignée végétale, classés parmi les procaryotes et les autres algues, possèdent également des pigments variés et des substances de réserve le plus souvent extraplastidiales, constituées de résidus glucane, mais dont les liaisons sont d’un type différent de celui qui caractérise l’amidon. La nature des substances de réserve et des pigments constitue un critère important de classification des protoctistes.
Ces pigments augmentent l’efficacité photosynthétique (surtout des plantes aquatiques) en absorbant dans une autre partie du spectre que la chlorophylle a.
Les chlorophylles a et b sont aussi présentes chez les Euglénophytes autotrophes (chez qui la présence de chloroplastes s’explique probablement par l’ingestion d’algues vertes par des individus hétérotrophes) ainsi que donc, chez les Chlorophycées. Ces caractéristiques partagées par ces dernières avec les Embryophytes montrent bien la parenté entre les algues vertes et les plantes terrestres.
 Figure 9
Figure 9
: spectre d’absorption lumineuse des chlorophylles a et b ainsi que des caroténoïdes
8. Les adaptations à la vie terrestre
Le milieu aquatique est un milieu homogène où les variations de température sont faibles. Le milieu terrestre est tout autre. Le sol et l’atmosphère sont des nouveaux espaces où la plante a dû aller chercher ses ressources. Les nouvelles contraintes environnementales ont engendré l’émergence d’organes spécialisés chez les plantes terrestres, comme les racines ou les feuilles.
Les premières plantes présentant un xylème comme organe vasculaire et de soutien, produisent encore une structure peu différenciée. C’est le cas du genre Cooksonia, le plus ancien fossile (datant de la fin du Silurien), unanimement actuellement considérée comme l’ancêtre des Trachéophytes. Chez cette espèce, dont les fossiles les plus anciens (environ 420 millions d’années) et classée dans la classe des Rhyniopsida, il n’y a pas de racines et de feuilles bien différenciées. Ces organes apparaîtront plus tard durant le Carbonifère chez les premières Lycopodophyta (Photo 5).
 Photo 5 : Lycopodium clavatum, une Lycopside actuelle aux nombreuses feuilles simples et petites
Photo 5 : Lycopodium clavatum, une Lycopside actuelle aux nombreuses feuilles simples et petites
Les plantes, sur le sol ferme, ont dû affronter deux problèmes majeurs : le dessèchement et la rétention des éléments nutritifs dans le sol.
Pour éviter une évaporation létale, les parties aériennes des plantes sont généralement couvertes d’une cuticule cireuse (Photo 6) qui leur permet de limiter les pertes en eau. Les échanges gazeux (CO2, oxygène) sont possibles grâce à de microperforations de l’épiderme (par exemple, chez les hépatiques à thalle, où les stomates sont absents et remplacés par des pores) qui évolueront rapidement en structures stomatiques. Également, pour lutter contre les UV, les plantes ont synthétisé des pigments protecteurs, comme les anthocyanes (Photo 7). Photo 6 : Coupe d’une feuille de narcisse avec l’épiderme.
Photo 6 : Coupe d’une feuille de narcisse avec l’épiderme.
 Photo 7 : Les tiges du géranium herbe-à-Robert prennent, due aux anthocyanes, une coloration plus rouge au soleil.
Photo 7 : Les tiges du géranium herbe-à-Robert prennent, due aux anthocyanes, une coloration plus rouge au soleil.
Mais, aussi, l’évolution des plantes terrestres est étroitement liée à un enjeu primordial, la gestion de l’eau. Celle-ci est indispensable au fonctionnement des cellules et à la survie de la plupart des organismes que nous connaissons. L’eau, avec les éléments vitaux qu’elle contient, doit donc être absorbée par la plante et stockée. Les racines, avec leurs nombreux poils absorbants, et le phénomène d’évapotranspiration permettent aux plantes terrestres de faire circuler l’eau du sol vers les organes photosynthétiques où se fabriquent la sève élaborée.
Et pour pouvoir accéder, dans le sol, à tous les éléments nutritifs dont elles ont besoin, les plantes ont, dès les premières installations sur la terre ferme, amorcé des relations avec les hyphes de champignons avec lesquels se sont formés des mycorhizes.
Si les relations symbiotiques avec des champignons supérieurs (comme les bolet, les amanites, les lactaires, etc) (Photo 8) sont bien connues pour de nombreux arbres forestiers, d’autres champignons mycorhiziens endogés vivent en étroite relation avec la plupart des plantes herbacées ou ligneuses de nos régions (comme des régions tropicales). Ces champignons font partie de la classe des Gloméromycètes (Photo 9), qui regroupent des mycètes (sans reproduction sexuée connue) dont des fossiles ont été trouvés déjà au Silurien à l’époque d’installation des plantes sur le continent. Dans cet échange mutualiste, le symbiote fongique reçoit des hydrates de carbone provenant de la plante et fonctionne comme un système racinaire étendu, améliorant ainsi considérablement l’absorption minérale par les racines.
 Photo 8 : Tricholoma cingulatum, un basiodiomycète symbiotique du saule.
Photo 8 : Tricholoma cingulatum, un basiodiomycète symbiotique du saule.
 Photo 9 : Glomus macrocarpus, une « grande » espèce de Gloméromycètes (quelques mm).
Photo 9 : Glomus macrocarpus, une « grande » espèce de Gloméromycètes (quelques mm).
Une autre caractéristique importante réside dans la capacité de produire la sporopollénine (le matériel organique d’origine biologique le plus résistant aux dégradations microbiologiques, chimiques et physiques) qui imprègne les parois des spores et des grains de pollen (comme son nom l’indique). Cette substance confère une grande résistance à des structures qui, après libération par la plante, seront confrontées aux aléas de l’environnement. La paléopalynologie, science des pollens fossiles, a pu mettre au jour des grains de sporopollénine encore intacts dans des roches sédimentaires datées de 470 millions d’années. Le cladogramme au chapitre « Origine des plantes » montre bien que l’occupation des milieux terrestres coïncide avec l’apparition de ce biopolymère.
Sa présence chez les Charophytes est signalée au niveau de la paroi du zygote.
 Photo 10: Grain de pollen de pin, muni de ses deux sacs aériens typique des conifères.
Photo 10: Grain de pollen de pin, muni de ses deux sacs aériens typique des conifères.
 Photo 11 : Sores (= ensemble de sporanges où sont produites les spores) chez Asplenium trichomanes.
Photo 11 : Sores (= ensemble de sporanges où sont produites les spores) chez Asplenium trichomanes.