
2. Les mousses (exemple du polytric commun)
A. Le gamétophyte et la reproduction sexuée
Le gamétophyte est issu de la méiospore haploïde. Celle-ci germe lorsque les conditions d’environnement sont favorables en produisant un réseau de filaments qui se ramifient abondamment et se différencient en deux types cellulaires distincts :
- un protonéma chlorophyllien à la surface du substrat qui évoque une algue verte dont les cloisons transversales seraient obliques et
- des rhizoïdes, avec peu ou pas de chloroplastes, qui s’enfoncent dans le substrat.
De place en place, le protonéma chlorophyllien produit, par recloisonnements, un bourgeon qui donnera naissance à un axe feuillé de 10 à 15 cm de haut, fixé au substrat par de nombreux rhizoïdes formant parfois un manchon feutré dense.
L’anatomie de la tige feuillée est simple (Figure 1). En coupe transversale on observe un épiderme sans stomates, un manchon cortical de cellules parenchymateuses plus ou moins allongées selon le grand axe et représentant, chez ces mousses plus évoluées, un faisceau conducteur élémentaire.
Celui-ci assure une certaine circulation de l’eau mais a surtout un rôle de soutien car l’essentiel de l’irrigation de la tige feuillée se réalise par un processus de capillarité, possible grâce à l’étroit agencement des phyllidies sur l’axe.
Les phyllidies sont finement dentées et leur nervure porte, à leur surface du sommet à la base, des lamelles uniassisiales dressées de 5 à 10 cellules de haut. Le cordon conducteur des phyllidies est normalement composé d’hydroïdes et de leptoïdes.
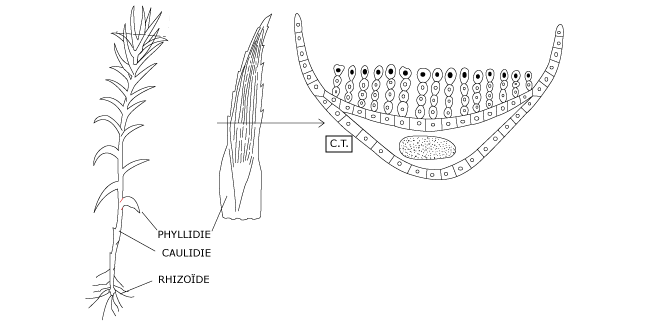 Figure 1 : Anatomie de la tige feuillée chez le polytric commun
Figure 1 : Anatomie de la tige feuillée chez le polytric commun
Pour des raisons qui restent à définir, une collerette de phyllidies parfois vivement colorées se forme au sommet des caulidies et, en son centre, apparaissent les gamétanges mâles (ou anthéridies) et les gamétanges femelles (ou archégones), soit sur un même pied (bisexué, mmonoécie), soit sur des pieds différents (unisexués, dioécie). Les gamétanges sont généralement pourvus d’organes effilés stériles, les paraphyses (Photo 2), qui jouent un rôle protecteur contre la dessiccation en favorisant la rétention d’eau par capillarité. Et sont entourées de feuilles souvent transformées (des bractées ou feuilles périchétiales)
L’anthéridie (Photo 1, Figure 2), brièvement pédicellée, est plus ou moins fusiforme et limitée par une paroi formée d’une seule assise cellulaire. Elle renferme un massif de cellules qui se transforment chacune en un anthérozoïde (gamète mâle) filiforme et plus ou moins spiralé, muni de deux flagelles. A maturité, l’anthéridie s’ouvre au sommet par gélification permettant la libération des anthérozoïdes qui, grâce à leurs flagelles, se déplacent dans le film d’eau qui recouvre les pieds de mousse.
 Photo 1 : Anthéridies entourées de paraphyses à l’extrémité d’une caulidie
Photo 1 : Anthéridies entourées de paraphyses à l’extrémité d’une caulidie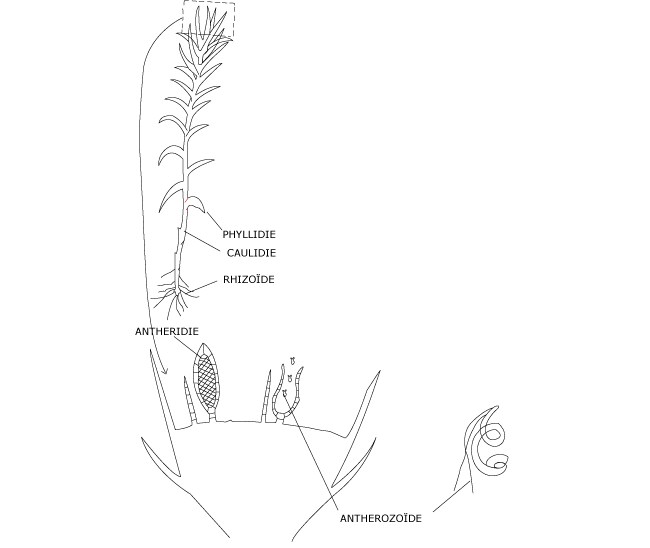 Figure 2 : Cycle de développement du polytric commun. Structure des gamétanges mâles : les anthéridies
Figure 2 : Cycle de développement du polytric commun. Structure des gamétanges mâles : les anthéridies
L’archégone (Figure 3), limité lui aussi par une paroi uniassisiale, revêt la forme d’une bouteille à col allongé dont la partie centrale, le canal du col, est obturée par une file de cellules. A maturité, les cellules du col dégénèrent en formant un mucilage qui gonfle en présence d’eau, provoquant l’ouverture du col et libérant l’accès pour les gamètes mâles tout en émettant une substance attirante pour les anthérozoïdes. Au fond de l’archégone, se différencie une cellule volumineuse : le gamète femelle ou oosphère.
La fécondation se réalise lorsque les tiges feuillées sont imbibées d’eau dans laquelle se meuvent les anthérozoïdes. Ceux-ci, attirés par le mucilage du col, atteignent l’oosphère par le canal de l’archégone
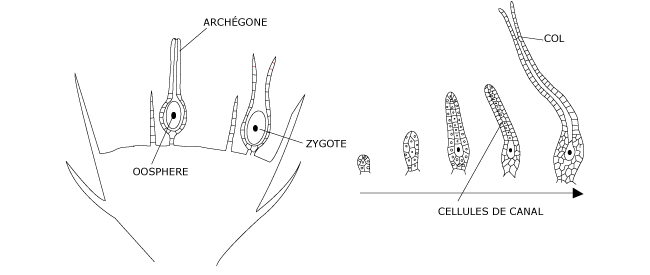 Figure 3 : Cycle de développement du Polytric commun. Structure des gamétanges femelles : les archégones
Figure 3 : Cycle de développement du Polytric commun. Structure des gamétanges femelles : les archégones
B. Le sporophyte et la reproduction agame
L’oeuf fécondé se cloisonne en un massif cylindrique. Vers le bas, il forme un pied qui s’enfonce dans le sommet de la caulidie et s’y fixe fermement. Vers le haut, il s’allonge en une sorte de hampe filiforme qui deviendra la soie (Figure 4).
Sous l’effet de cette élongation, la jeune soie déchire le sommet et le col de l’archégone qui, continuant leur croissance, forment une coiffe protégeant le sporange en formation. En effet, l’extrémité distale de la soie se renfle et s’organise pour devenir un sporange (= capsule ou sporogone). A maturité, le sporange, dont la croissance s’est faite aux dépens du parenchyme lacuneux, est devenu un “sac” traversé par la columelle persistante. La coiffe tombe, un opercule apical se détache au niveau d’une fente annulaire de déhiscence et le sporange s’ouvre en découpant une dernière paroi protectrice en une (deux chez d’autres genres) couronne de dents, le péristome (parfois accompagné d’un épiphragme, e.a. chez le Polytric), qui peuvent se rabattre ou se redresser selon l’état hygrométrique de l’air. Souvent, la soie s’infléchit et les spores sont libérées et disséminées (Photo 2).
 Figure 4 : Cycle de développement du Polytric commun. Germination des spores en protonéma, cycle et structure du sporophyte
Figure 4 : Cycle de développement du Polytric commun. Germination des spores en protonéma, cycle et structure du sporophyte
 Photo 2 : Sporophytes du polytric avec et sans coiffes
Photo 2 : Sporophytes du polytric avec et sans coiffes
En coupe transversale, un sporange jeune (Figure 5) présente successivement, de l’extérieur vers l’intérieur :
- une assise externe qui constitue un épiderme percé de stomates plus ou moins rudimentaires,
- un parenchyme lacuneux, chlorophyllien même en profondeur, interrompu par des lacunes en files axiales,
- un tissu dense qui forme un manchon d’archéspores,
- un cylindre axial massif, gorgé d’eau, qui constitue la columelle.
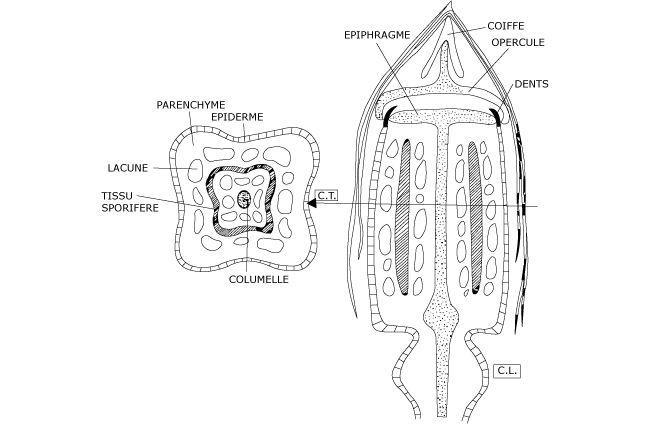 Figure 5 : Structure du sporange du Polytric commun
Figure 5 : Structure du sporange du Polytric commun
La sporogenèse se déroule selon le processus indiqué à la figure 6.
 Figure 6 : Sporogénèse
Figure 6 : Sporogénèse
C. Le cycle de développement complet du Polytric
Le cycle montre la succession des deux formes :
- le gamétophyte constitué par le protonéma (souvent éphémère), puis les axes feuillés
- le sporophyte constitué du sporogone qui se développe en restant fixé sur le gamétophyte. Le gamétophyte représente la phase haploïde et le sporophyte représente la phase diploïde (voir figure 7).
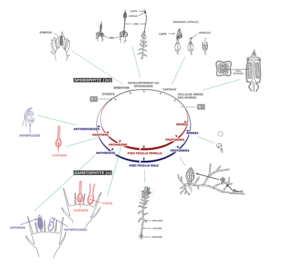 Figure 7 : Le cycle de développement du Polytric, mousse
Figure 7 : Le cycle de développement du Polytric, mousse

3. Les hépatiques (exemple de Marchantia polymorpha)
A. Le gamétophyte et la reproduction sexuée (voir figure 8)
Les méiospores germent pour donner un protonéma haploïde très court à partir duquel une lame verte foliacée, appliquée sur le substrat, prend rapidement naissance. Le gamétophyte adulte peut être considéré comme un thalle. Il est édifié par quelques cellules méristématiques situées dans l’échancrure apicale du thalle (et non par une cellule apicale unique comme chez le Polytric) et se ramifie de façon dichotomique. Sa face supérieure est marquée de losanges réguliers percés chacun d’un pore central limité par des cellules de garde. Celles-ci forment une espèce de tonnelet de hauteur supérieure à l’épaisseur de l’épiderme : il déborde donc celui-ci au-dessus et en dessous. Sous l’épiderme se trouvent des chambres aérifères, chacune d’elles correspondant à un des losanges repérés de l’extérieur. Ces chambres sont délimitées par des cloisons verticales de 3 à 4 cellules riches en chloroplastes. Chacune de ces chambres représente ainsi une cavité assimilatrice et également facilite la respiration. Sous le plancher des cavités assimilatrices, le parenchyme à grandes cellules est un parenchyme de réserve avec cellules oléagineuses et mucilagineuses.
Le thalle est encore parcouru par une fausse nervure médiane visible en surface et formée de cellules allongées selon le grand axe de l’organe.
La face inférieure du thalle est pourvue de rhizoïdes et d’écailles imbriquées sur deux rangs, une de chaque côté de la nervure. Ces écailles peuvent être interprétées comme représentant des appendices foliaires réduits.
Durant la bonne saison, les thalles en croissance produisent des rameaux spécialisés, unisexués, dressés (+/- 3 cm de haut), insérés dans le prolongement direct de la nervure, au point où fonctionnait le méristème : ce sont les gamétangiophores (Photo 1).
 Photo 4 : Marchantia polymorpha avec des gamétangiophores femelles
Photo 4 : Marchantia polymorpha avec des gamétangiophores femelles
Les rameaux produits par certains thalles – qui s’avéreront être des thalles mâles – portent, au sommet d’un axe, un plateau lobé dont l’anatomie est identique à celle du thalle. Néanmoins, entre les chambres aérifères, se différencient des anthéridies enfoncées dans des cavités ovoïdes formées par des replis de l’épiderme; ces cavités s’ouvrent par un petit canal à la face supérieure. L’éclatement des anthéridies libère les anthérozoïdes qui sont projetés sur le plateau dans un film d’eau.
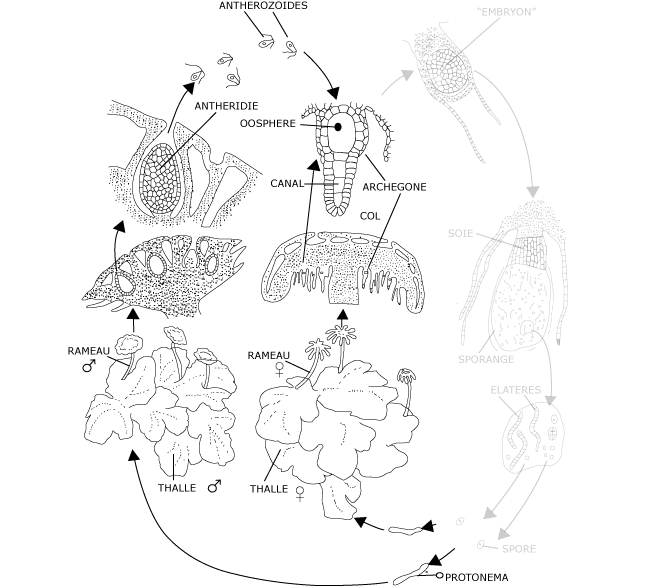 Figure 8: Le cycle de développement du Marchantia : gamétophyte et reproduction sexuée (la partie droite en grisée reprend le sporophyte et la reproduction agame, détaillée ci-après – part B)
Figure 8: Le cycle de développement du Marchantia : gamétophyte et reproduction sexuée (la partie droite en grisée reprend le sporophyte et la reproduction agame, détaillée ci-après – part B)
Les rameaux spécialisés produits par les thalles femelles ont une structure analogue à celle des rameaux mâles. Ils se distinguent cependant par leurs lobes plus profondément échancrés, généralement au nombre de 9. Les archégones, au nombre de 12 à 16 et de structure fort semblable à celle des archégones de polytric, sont initiés entre les lobes de manière centripète, c’est-à-dire que les premiers à être formés se situent le plus loin de l’axe de l’archégoniophore et les suivants en sont de plus en plus près. Ils sont protégés par un involucre (sorte de collerette) tendu entre deux lobes contigus. Le col des archégones est haut de 13 cellules ; le canal n’en possède que quatre.
La fécondation a lieu par temps humide, avant l’élongation complète du rameau spécialisé. Elle se produit grâce au film d’eau qui recouvre les thalles.
L’œuf fécondé (zygote) se développe à l’intérieur de l’archégone dont les parois se modifient pour abriter le jeune sporophyte en croissance.
Il existe aussi une reproduction asexuée. Des propagules sont produites dans des corbeilles sur la face supérieure du thalle.

Photo 5 : Marchantia polymorpha avec corbeilles à propagules
B. Le sporophyte et la reproduction agame (voir figure ci dessous)
Le sporophyte est nettement réduit par rapport à celui des mousses. La soie – l’axe portant le sporange – fragile reste très courte et ce n’est que peu avant la maturité que le sporange est poussé en avant et déchire le sommet de l’archégone accrescent. Le sporange ne possède ni columelle, ni péristome.
Une coiffe évanescente persiste, à la base de la soie, sous forme d’un petit manchon. La capsule s’ouvre par 4 à 6 valves qui sont des déchirures de la paroi plutôt que des fentes de déhiscence. Les spores sont accompagnées de structures sensibles à l’hygrométrie qui facilitent leur dispersion et la dissémination : les élatères. Celles-ci se différencient, avant la formation des archéspores, par division au sein des tissus du sporange. Elles seraient également impliquées dans la poussée qui fait éclater le sporange.
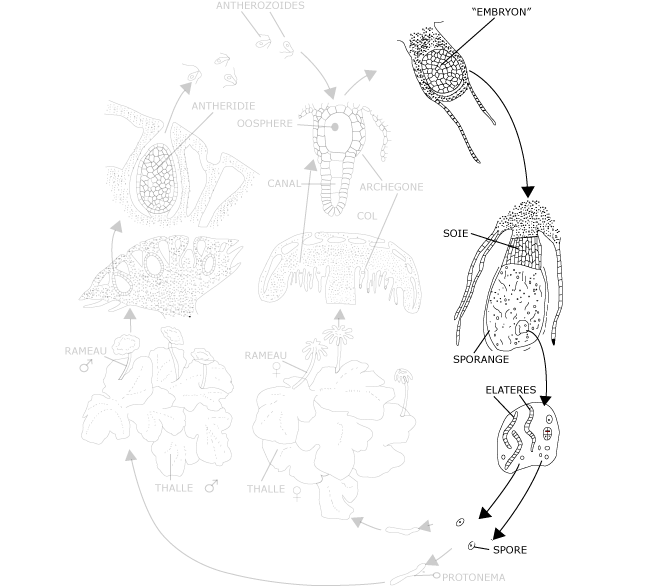 Figure 10 : Le cycle de développement du Marchantia : sporophyte et reproduction agame
Figure 10 : Le cycle de développement du Marchantia : sporophyte et reproduction agame
Si ce n’est l’absence d’une structure dressée portant les gamétanges, les étapes jusqu’à l’embryon et les organes reproductifs sont assez similaires dans l’autre grande classe au sein des hépatiques (Jungermanniopsida).