
1. Les prêles
Les prêles appartiennent à un genre cosmopolite (Equisetum) et comptent une quinzaine d’espèces. Lorsqu’on remonte l’échelle du temps, on constate que le type “prêle” est présent depuis la fin du Carbonifère (> 280 millions d’années) ; le genre Equisetum n’est cependant connu avec certitude que depuis le Crétacé. Depuis, il a été en s’amenuisant quelque peu. Il est proche parent d’un groupe, entièrement disparu depuis le Triasique, mais qui a occupé une place prépondérante dans les végétations dès le Dévonien supérieur, soit durant près de 150 millions d’années. Mis à part les Psilophytes, il représente probablement le type le plus ancien de plante vasculaire de la flore actuelle. L’exemple choisi est celui de la prêle des champs (Equisetum arvense) (Photo 1).
 Photo 1 : Tiges fertiles de la prêle des champs (avril-mai)
Photo 1 : Tiges fertiles de la prêle des champs (avril-mai)
A. Reproduction agame
A.1 Origine et structure des sporanges
Certains rameaux aériens, très précoces, non chlorophylliens et non ramifiés, portent des sporanges réunis en un cône (Photo 2) ; ce strobile (= cône à écailles membraneuses pour l’Equisetum) possède donc un axe prolongeant celui de l’axe aérien et porte de courtes ramifications se terminant par une plaquette polygonale, le sporangiophore.
 Photo 2 : Strobile d’Equisetum
Photo 2 : Strobile d’Equisetum
Lorsque le strobile est jeune, ces plaquettes vues de l’extérieur sont étroitement agencées formant une espèce de carrelage; à maturité, elles s’écartent et permettent la libération des spores car, sur la face interne de chacune de ces plaquettes sont apparues entre temps ,une dizaine de sporanges (Figure 1).
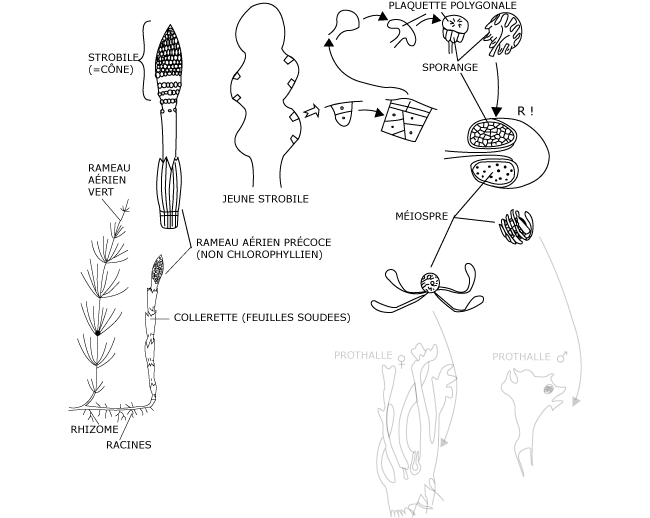 Figure 1 : Cycle de reproduction de la prêle des champs. Sporophyte et reproduction agame
Figure 1 : Cycle de reproduction de la prêle des champs. Sporophyte et reproduction agame
Chaque sporange provient du fonctionnement d’une seule cellule superficielle qui se cloisonne de façon péricline ; la cellule externe produit les assises de la paroi sporangiale, tandis que l’interne donne les archéspores (= cellules-mères de spores).
Les assises de paroi, au nombre de quatre à cinq, se réduisent à deux à maturité; elles comportent des épaississements spiralés intervenant dans la déhiscence; le tapis provient des cellules externes aux archéspores.
A.2 Les spores
La méiose est normale et produit des méiospores en apparence toutes semblables. En plus de l’intine et de l’exine, les spores forment extérieurement une troisième paroi qui, à maturité, se découpe en lanières qui demeurent attachées en un point de l’exine et sont élargies en spatule à leur extrémité. Enroulées en spirale autour de la spore par forte humidité atmosphérique, ces élatères (Photo 3) se déploient et s’étalent dans l’air sec.
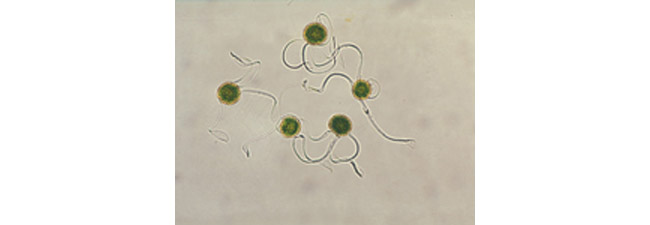 Photo 3 : Spores avec élatères d’Equisetum
Photo 3 : Spores avec élatères d’Equisetum
Les mouvements brusques de détente de ces lanières ont pour effet vraisemblable de favoriser la dissémination alors que les inévitables accrochages qu’elles peuvent avoir entre elles ont un effet inverse, celui de maintenir groupées les spores qui, en germant, donneront des prothalles unisexués.
B. La reproduction sexuée
B.1 Le gamétophyte (Figure 2)
Les spores germent dès qu’elles ont atteint le sol et forment des prothalles discrets, fugaces et aériens. Ils sont composés de lobes laciniés qui leur confèrent une forme très irrégulière.
Au bout d’un mois environ, les gamétanges naissent d’une cellule superficielle qui grossit et se développe vers l’intérieur en se cloisonnant.
Le prothalle femelle est légèrement plus grand que le gamétophyte mâle; il est également plus découpé en languettes chlorophylliennes stériles, les archégones étant confinés à la partie massive.
Lors de la différenciation de l’archégone, après la première division de l’initiale, la cellule externe donne, par une division anticline, un col assez court, tandis que la cellule profonde se cloisonne de manière péricline pour donner deux cellules de canal, une cellule ventrale de canal, une oosphère et une cellule de base.
Le prothalle mâle, moins découpé porte également ses gamétanges (les anthéridies) sur le lobe principal. L’anthéridie donne 256 anthérozoïdes pluriciliés et s’ouvre sous la pression interne par une petite ouverture triangulaire préalablement délimitée dans la paroi externe par trois cloisonnements anticlines.
B.2 Le zygote et l’embryon (Figure 2)
La fécondation a nécessairement lieu en période humide. Une fois l’oosphère fécondée, le zygote se cloisonne et sa première division est perpendiculaire au grand axe de l’archégone ; les deuxième et troisième divisions du jeune embryon se produisent suivant un axe orienté à 90° et donnent quatre quadrants inégaux : des deux supérieurs, le plus grand donne la tige ou rhizome (T) et le plus petit, la première feuille (F) ; des deux inférieurs, le plus grand donne la première racine (R) et le plus petit, le pied (P). L’embryon, ainsi conformé, peut croître immédiatement.
 Figure 2 : Cycle de reproduction de la prêle des champs – Gamétophyte et reproduction sexuée.
Figure 2 : Cycle de reproduction de la prêle des champs – Gamétophyte et reproduction sexuée.
C. Cycle de reproduction de la prêle des champs

2. Les fougères
Les fougères présentent un type de reproduction sexuée fort voisin de celui des prêles, jusque dans les détails, bien que les deux groupes semblent avoir évolué séparément depuis plus de 300 millions d’années. L’appareil végétatif aérien paraît souvent plus développé chez les fougères (cfr. les fougères arborescentes), mais ce caractère n’a qu’une importance accessoire quand on sait que des Sphénophytes fossiles, tels les Calamites, pouvaient atteindre la taille d’arbres de 30 m de haut. On notera cependant que, pour la première fois au cours de l’évolution, l’appareil foliaire (nommé fronde) peut déployer des surfaces considérables (mégaphylles) grâce à un système de nervuration en nombreux cordons vasculaires anastomosés qui prennent leur origine au niveau du cylindre central de la tige où des fenêtres foliaires sont ménagées comme chez les plantes supérieures. Il s’agit donc d’un caractère évolué. L’exemple sur lequel s’appuiera la démonstration est celui de la fougère mâle (Dryopteris filix-mas) (Photo 4).
 Photo 4 : Dryopteris filix-mas
Photo 4 : Dryopteris filix-mas
A. La reproduction agame
A.1 Origine et structure des sporanges
Les sporanges se forment à la face inférieure des frondes, de part et d’autre des nervures secondaires. Ils sont groupés en sores (Photos 4, 5, 6), abrités sous un repli de l’épiderme, l’indusie (voir Figure 4).
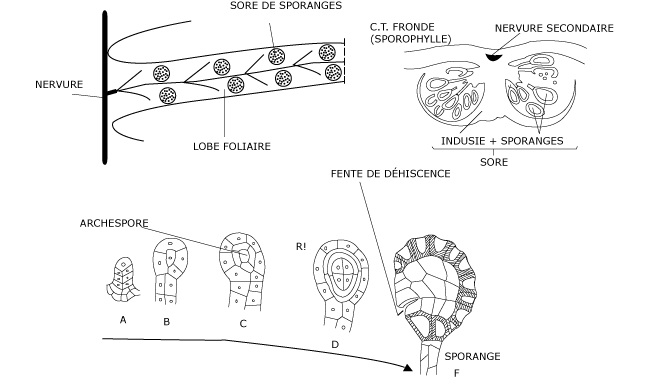 Figure 4 : Structure des sores et des sporanges
Figure 4 : Structure des sores et des sporanges
N.B. : L’organisation, les modalités de répartition des sporanges et des sores et le degré de protection de ceux-ci présentent une très grande diversité chez les Fougères.
 Photo 4: Sores de fougère scolopendre
Photo 4: Sores de fougère scolopendre
 Photo 5 : Sores avec indusie (protection)
Photo 5 : Sores avec indusie (protection)
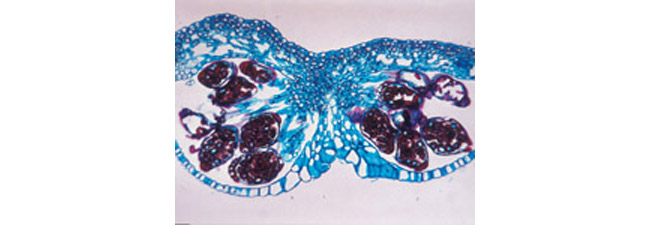 Photo 6 : Sore (en gros plan) avec sporanges (parties foncées) à la face inférieure de la fronde de la fougère mâle
Photo 6 : Sore (en gros plan) avec sporanges (parties foncées) à la face inférieure de la fronde de la fougère mâle
Chaque sporange se différencie à partir d’une cellule épidermique et se cloisonne dans un ordre précis pour former un organe lenticulaire porté par un axe plus ou moins allongé. Cet organe comporte une assise périphérique et, au début, une seule cellule centrale de forme pyramidale. Celle-ci se cloisonne encore sur ses quatre faces en délimitant une nouvelle assise périphérique qui s’organise en tapis nourricier.
L’unique cellule centrale du jeune sporange est une archéspore qui subit une sporogenèse normale (voir Figure 5) et produit finalement des méiospores à triple paroi.
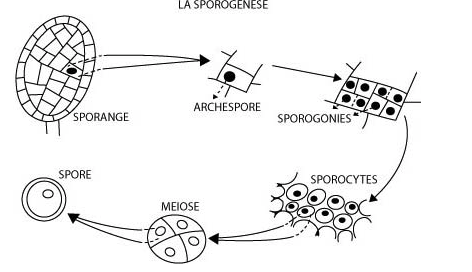 Figure 5 : Sporogénèse
Figure 5 : Sporogénèse
A.2 Déhiscence du sporange et dissémination des spores
Au cours de la maturation du sporange, une file verticale de cellules de l’assise externe se différencie en anneau mécanique : les parois cellulaires internes et anticlines de ces cellules s’épaississent considérablement et se lignifient, dessinant un U dont l’ouverture est tournée vers l’extérieur.
A maturité, le sporange se dessèche ; l’évaporation progressive de l’eau des cellules de l’anneau mécanique, au travers de la paroi externe demeurée mince, provoque une réduction du volume cellulaire entraînant, vers l’extérieur, la paroi amincie qui constitue une zone de moindre résistance.
Les deux branches du U ne pouvant se rapprocher, car leur paroi radiale est rigide, une traction tangentielle se communique jusqu’à l’extrémité de l’anneau mécanique où les cellules à parois non épaissies se rompent, tandis que l’arc des cellules différenciées reprend sa liberté.
Grâce à ce mécanisme très efficace, le sporange tout entier s’ouvre donc par une large fente transversale projetant à l’extérieur les spores mûres.
 Figure 6 : Déhiscence du sporange et dissémination des spores
Figure 6 : Déhiscence du sporange et dissémination des spores
B. La reproduction sexuée
B.1 Le gamétophyte
En germant, la spore se cloisonne et produit un thalle formé d’une lame foliacée, chlorophyllienne comportant une à plusieurs assises cellulaires (Figure 7). C’est le prothalle, dont la taille ne dépasse guère 1 cm de largeur et dont la forme est plus ou moins cordée.
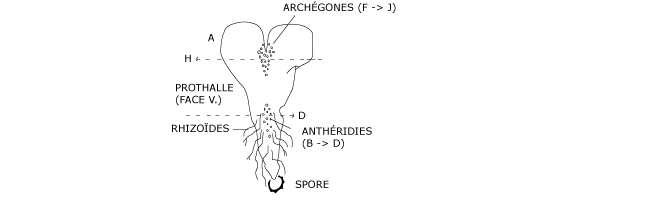 Figure 7 : Formation d’un prothalle à partir d’une spore de fougère mâle.
Figure 7 : Formation d’un prothalle à partir d’une spore de fougère mâle.
Ce prothalle croît grâce au fonctionnement d’une cellule initiale, logée entre les deux lobes. Appliqué sur le sol, il y est fixé par des rhizoïdes unicellulaires. La lame prothallienne est pluriassisiale dans sa partie centrale. Les gamétanges se forment à la face ventrale du prothalle (Figures 8 & 9), contre le substrat par conséquent.
Anthéridies et archégones apparaissent sur le même prothalle mais il arrive que les anthéridies soient mûres longtemps avant les archégones, ce qui favorise la fécondation croisée.
Les anthéridies (Figure 8), assez rudimentaires, s’ouvrent et libèrent des anthérozoïdes plus ou moins spiralés, pourvus d’une couronne de nombreux cils.
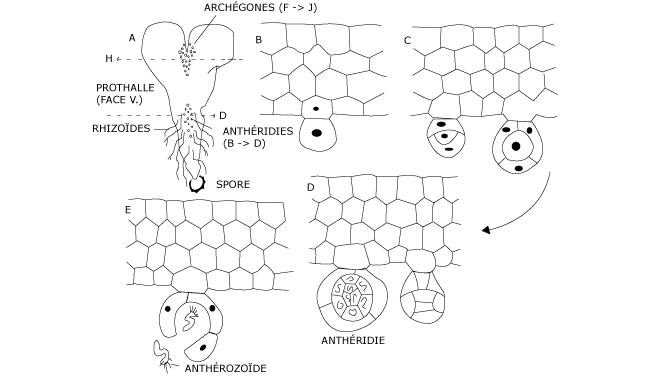 Figure 8 : Formation des anthéridies et libération des anthérozoïdes
Figure 8 : Formation des anthéridies et libération des anthérozoïdes
Avec un col fort court dirigé vers le bas, les archégones (Figure 9) – assez simples aussi – renferment une oosphère et quelques cellules de canal seulement.
 Figure 9 : Formation des archégones
Figure 9 : Formation des archégones
B.2 Fécondation et développement du zygote
Comme chez les Bryophytes, les anthérozoïdes libérés nagent dans le film d’eau humectant le sol en surface ; ils sont attirés vers les archégones par chimiotactisme.
Les deux premières divisions du zygote le partagent en quatre quadrants qui, par recloisonnements, vont fournir des organes différents : un pied (P) massif qui rattache l’embryon en développement aux tissus du prothalle dont il se nourrit au début ; une racine (R) qui pénètre dans le sol ; une tige (T) et une première fronde (= feuilles) (F) qui, dans sa croissance, se charge de chlorophylle.
Comme chez les Prêles, il y a donc production d’un authentique embryon. Bien que plusieurs archégones se forment sur le même prothalle, un seul embryon poursuit normalement sa destinée. Dès que les jeunes frondes sont épanouies, le prothalle disparaît.
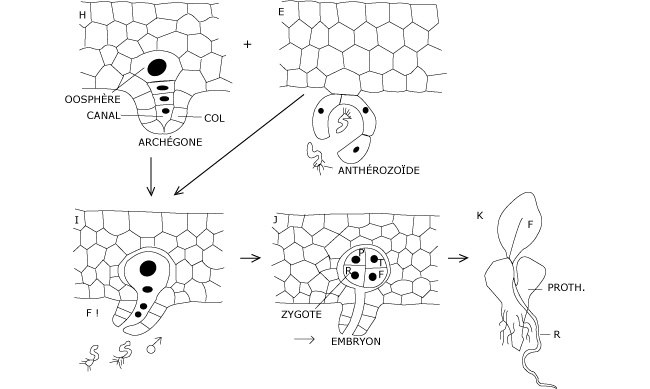 Figure 10 : Fécondation et développement de l’embryon
Figure 10 : Fécondation et développement de l’embryon
C. Cycle de reproduction de la fougère mâle
 Figure 11 : Cycle de reproduction de la fougère mâle
Figure 11 : Cycle de reproduction de la fougère mâle
3. Les sélaginelles
La formation de deux types de méiospores chez une même espèce constitue une acquisition significative pour les végétaux : il s’agit d’un trait caractéristique et « définitif » de l’évolution végétale. Un des premiers groupes à présenter cette caractéristique est celui des Lycophytes ; cependant, elle apparaît également chez d’autres lignées de Ptéridophytes, notamment chez des fougères, du moins les fougères aquatiques (Hydroptéridales).
Nous n’envisageons qu’un seul cas, celui des Sélaginelles (Photo 7) qui font partie des Lycophytes.
 Photo 7 : Sélaginelle sp
Photo 7 : Sélaginelle sp
A. La reproduction agame
A l’extrémité de certains rameaux dressés, toutes les feuilles se transforment en sporophylles : feuilles plus larges et plus serrées les unes contre les autres et qui portent à leur aisselle des sporanges qui sont donc groupés en épis, les strobiles (Figure 12).
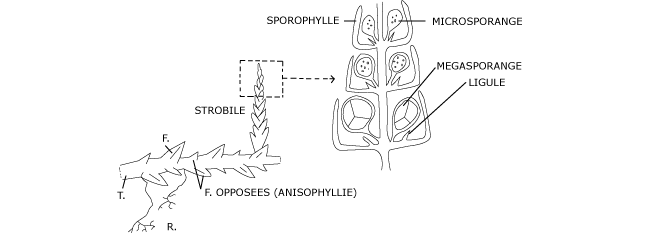 Figure 12 : Sporophyte, strobile et sporange de sélaginelle
Figure 12 : Sporophyte, strobile et sporange de sélaginelle
En réalité il se forme deux sortes de sporanges au sein des strobiles : à la base, souvent, des mégasporanges de plus grande taille que les microsporanges (Figure 4) situés plus haut sur l’axe.
Dans le microsporange, se réalise une sporogenèse normale qui aboutit à la formation de nombreuses microspores.
Par contre, dans le mégasporange, toutes les sporogonies dégénèrent, à l’exception d’une seule qui devient l’unique sporocyte. Il n’y a donc que quatre mégaspores qui sont formées. Elles sont volumineuses, jusqu’à plus de 20 fois plus grosses que les microspores, riches en substances de réserve et entourées d’une paroi très épaisse. Il arrive même qu’une seule mégaspore privilégiée persiste.
Tous les sporanges s’ouvrent à maturité et libèrent les spores (Figure 13). Cette dissémination peut être différée : la mégaspore privilégiée peut en effet poursuivre son développement sur la plante-mère.
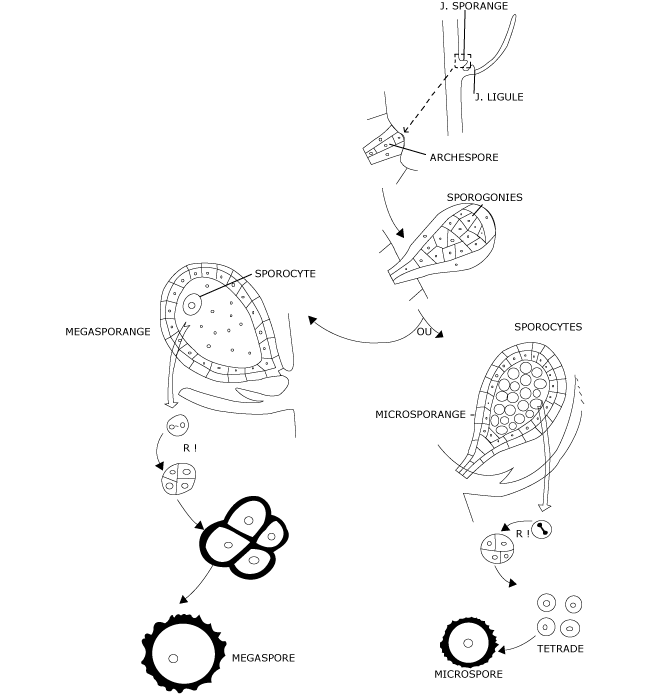 Figure 13 : Mégasporogenèse et microsporogenèse au sein du strobile
Figure 13 : Mégasporogenèse et microsporogenèse au sein du strobile
B. La reproduction sexuée
B.1 Le gamétophyte mâle
La microspore se divise en formant une petite cellule dite végétative qui, à elle seule, représente tout le prothalle et une grande cellule qui se cloisonne de manière à donner une paroi uniassisiale et des cellules centrales qui évoluent en anthérozoïdes biciliés. C’est donc une anthéridie rudimentaire qui s’ouvre et libère les gamètes mâles.
B.2 Le gamétophyte femelle
Dans la mégaspore, le noyau se divise plusieurs fois successivement et les noyaux produits se dispersent dans la masse cytoplasmique. Alors seulement intervient un cloisonnement différentiel qui partage le massif cellulaire en deux régions : un « tissu » inférieur, formé de grandes cellules où s’accumulent des substances de réserve, et un tissu « supérieur » beaucoup plus dense car il est formé de petites cellules ; ce tissu, en s’hydratant, exerce une pression sur l’épaisse paroi mégasporale qui se rompt. En surface, sous l’ouverture pariétale de plus en plus élargie, se forment les archégones rudimentaires.
Tout ce développement prothallien s’accomplit normalement au sol, après dissémination de la mégaspore, mais il peut arriver qu’il débute et même se réalise entièrement sur le sporophyte lui-même.
B.3 Fécondation et formation de l’embryon
L’oopshère est fécondée par la pénétration de l’anthérozoïde, mobile dans l’eau. Habituellement, une seule oopshère est fécondée et poursuit son développement dans le prothalle. Le zygote se divise d’abord pour donner un « suspenseur » qui entraîne l’embryon dans les tissus nourriciers profonds. L’embryon se différencie comme chez les Fougères. Finalement, la jeune plantule se dégage de l’enveloppe mégasporale et reconstitue un sporophyte.
 Figure 12 : Les gamétophytes, les gamétanges et la fécondation chez la sélaginelle
Figure 12 : Les gamétophytes, les gamétanges et la fécondation chez la sélaginelle
C. Cycle de reproduction de la sélaginelle
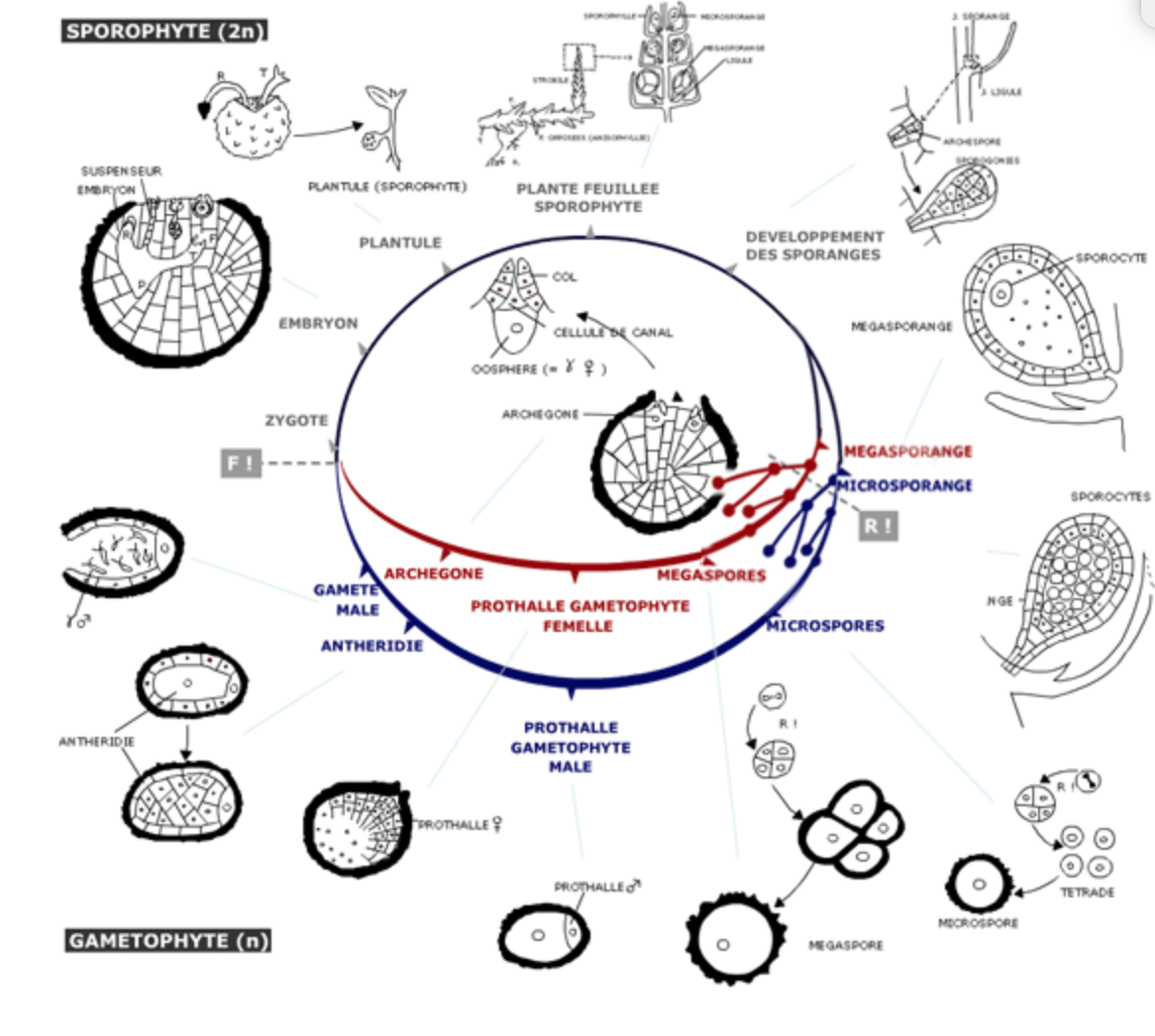 Figure 13 : Cycle de reproduction de la sélaginelle
Figure 13 : Cycle de reproduction de la sélaginelle