
Outre la prédominance du sporophyte qui constitue un caractère définitivement acquis dans la lignée des végétaux, outre aussi l’hétérosporie qui sera désormais observée chez tous les groupes considérés comme « plus avancés », l’évolution a également porté sur l’appareil végétatif.
L’adaptation au milieu terrestre pose en effet de nombreux problèmes tant physiologiques que morphologiques, engendrés, d’une part, par l’éloignement des ressources hydriques et trophiques et d’autre part, par l’environnement desséchant dans lequel les axes aériens et leurs appendices, les feuilles, se déploient. Les caractères adaptatifs appropriés vis-à-vis de ces contraintes sont apparus progressivement au cours de l’évolution. Ils visent :
(1) à limiter les pertes en eau par les structures aériennes, avec l’apparition d’une cuticule (Photo 1) sur les surfaces en contact avec l’atmosphère et la différenciation concomitante des stomates. Cette adaptation limite la transpiration, mais laisse une voie d’accès privilégiée au CO2 essentiel pour assurer la photosynthèse.
N.B. : l’élaboration de l’enveloppe de sporopollénine autour des spores serait la première acquisition majeure ; cette innovation permet la dispersion de ces propagules dans des milieux secs.
(2) à assurer une circulation rapide de l’eau jusqu’aux extrémités des plus hautes ramifications aériennes. La capacité de produire la lignine, présente dans les parois des cellules des tissus conducteurs, constitue également une étape critique de l’évolution des plantes. La rigidité des Bryophytes était assurée par la seule turgescence des cellules, ce qui limitait leur possibilité de croissance en hauteur. La lignine contribue de manière marquante à la rigidité des parois du sporophyte des plantes vasculaires.
 Photo 1 : Feuilles à cuticule cireuse chez Populus trichocarpa
Photo 1 : Feuilles à cuticule cireuse chez Populus trichocarpa
(3) à optimaliser l’exploitation de ressources contenues dans le substrat ; les racines ne sont plus seulement des organes de fixation à ce substrat mais contribuent à l’approvisionnement en eau et en éléments minéraux.
Chez les Ptéridophytes tenus pour les plus anciens, le sporophyte est constitué par un système d’axes à ramifications dichotomes analogues à celles que l’on observe chez les thalles des algues ou des hépatiques ; on ne peut y reconnaître ni feuilles, ni racines, ni même de véritables tiges. De toute évidence, ces organes ne sont pas apparus brusquement, mais les documents paléontologiques ne permettent pas de suivre les étapes de cette évolution vers des organes bien différenciés. Différentes théories ont alors été proposées pour tenter de reconstituer l’origine de ces structures.
1. Origine de la tige feuillée
Deux théories ont été élaborées pour expliquer comment, à partir du thalle des premières plantes vasculaires primitives, la pousse feuillée s’était progressivement constituée. Elles peuvent être résumées de la manière suivante.
(1) L’organe originel serait un axe dépourvu de tout appendice tel qu’il peut s’observer sur les restes des lointains ancêtres des Ptéridophytes actuels. Les appendices apparaîtraient sous forme d’excroissances édifiées dans les tissus superficiels des axes nus ; ces bourrelets plus ou moins allongés seraient dans un premier temps dépourvus de toute vascularisation.
Ensuite, le cylindre conducteur de l’axe émettrait des cordons vasculaires qui gagneraient progressivement les appendices foliacés. Cette hypothèse fournit une interprétation plausible quant à l’origine des petits appendices foliacés (lignée des microsporophylles) tels qu’on les connaît chez de nombreux fossiles (Asteroxylon) mais aussi les Lycopodes et les Sélaginelles actuels.
 Figure 1 : Première théorie de l’origine de la tige feuillée chez les Ptéridophytes.
Figure 1 : Première théorie de l’origine de la tige feuillée chez les Ptéridophytes.
(2) Une deuxième théorie considère que c’est l’axe nu, ramifié de façon dichotomique, qui aurait donné tiges et feuilles de la manière suivante :
un axe principal se dégage grâce à l’asymétrie qui s’installe progressivement dans les dichotomies successives, les plus courtes (ou les plus faibles) étant rejetées latéralement. Ces axes courts se seraient ensuite disposés dans un même plan, au lieu de se disposer successivement dans des plans décalés de 90°. Enfin, les ramifications latérales disposées dans un même plan se seraient enveloppées d’une lame de tissu qui les réunirait pour former le limbe avec son réseau de nervures.
Cette hypothèse permet d’expliquer l’origine des grandes frondes (mégasporophylles) (Photo 2), mais aussi celle des petits appendices foliacés par réduction des ramifications latérales à un seul axe qui se transforme en feuille.
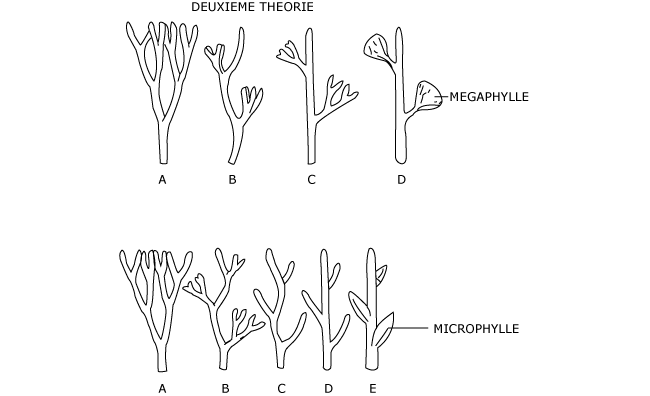 Figure 2 : Deuxième théorie de l’origine de la tige feuillée chez les Ptéridophytes
Figure 2 : Deuxième théorie de l’origine de la tige feuillée chez les Ptéridophytes
 Photo 2 : Une fronde frêle dans les lieux frais : Cystopteris fragilis .
Photo 2 : Une fronde frêle dans les lieux frais : Cystopteris fragilis .
3. Evolution du système conducteur
Chez les Thallophytes, les tissus conducteurs sont à peine ébauchés. Ainsi, chez la laminaire, la fronde du sporophyte montre, en coupe transversale, un cylindre central en périphérie duquel apparaissent des files longitudinales de cellules allongées, et manifestement conductrices, que l’on assimile à des tubes criblés. Il n’existe cependant aucune ébauche de cordon conducteur au sens où il a été défini chez les Angiospermes.
Chez les Bryophytes, l’axe aérien du gamétophyte montre bien un cylindre central de cellules allongées mais ne possédant qu’une différenciation histologique rudimentaire. Son rôle conducteur est certainement peu accusé. La lignine est totalement absente.
Chez les végétaux actuels, seuls les Ptéridophytes et les Spermatophytes possèdent de véritables vaisseaux. C’est pourquoi on les appelle également « Végétaux vasculaires » ou « Trachéophytes ».
Entre les structures conductrices des Ptéridophytes les plus simples et les plus évoluées des Thallophytes ou des Bryophytes, l’hiatus est cependant considérable. Comment donc seraient apparues ces structures conductrices essentielles pour assurer l’alimentation et l’irrigation d’un appareil végétatif se déployant à de grandes distances de la source en eau et en éléments minéraux ? Sont-elles apparues brusquement en même temps que la prédominance du sporophyte aérien ou, au contraire, cette transformation a-t-elle suivi un long cheminement ?
Dans cette dernière éventualité, les intermédiaires auraient donc disparu et seule l’étude des fossiles permettrait de reconstituer une lignée évolutive plausible.
1. Le concept de la stèle
La stèle (du latin stela, colonne) correspond à l’ensemble des structures conductrices ou, de façon plus large, au cylindre central composé principalement – il est vrai – de structures conductrices.
Ce concept est évident lorsqu’il est appliqué à la racine et même, chez les Dicotylédones à anneau conducteur continu, à la tige ;
il est plus flou chez les Dicotylédones à faisceaux conducteurs discrets situés sur un seul cercle et, plus encore, chez les tiges de Monocotylédones. Quoi qu’il en soit, ces dispositions reconnues chez les Angiospermes, ont été précédées dans le temps par d’autres structures que l’on interprète en conséquence comme plus primitives. Celles-ci peuvent être ordonnées et s’insèrent assez bien dans une série évolutive qui montre comment s’est formé le cylindre conducteur des plantes actuelles.
2. L’évolution de la stèle
1) Chez les Ptéridophytes considérés comme les plus primitifs, les tissus conducteurs de la tige, centraux et axiaux, forment un cylindre plein et continu, une sorte de colonne qui comporte un xylème endarche (ou xylème primaire occupant une position centrale) entouré d’un manchon de phloème (protostèle).
2) D’autres Ptéridophytes, moins primitifs, forment une stèle un peu différente, constituée de deux manchons emboîtés, le central étant xylémien, l’externe phloémien (siphonostèle). L’axe de la tige est occupé par une moelle.
3) Plusieurs fougères, tenues pour les plus évoluées parmi les Ptéridophytes, montrent la même structure, mais l’anneau conducteur est percé aux nœuds de fenêtres foliaires (dictyostèle). En coupe transversale, par conséquent, au niveau des brèches au moins, la stèle apparaît comme fragmentée en cordons conducteurs.
4) Enfin, cette évolution est la mieux marquée finalement chez les Spermatophytes, où les ouvertures foliaires sont notables, avec des cordons conducteurs bien individualisés et en continuité parfaite avec les traces foliaires (eustèle). Ces cordons se disposent plus ou moins en cercle (Dicotylédones herbacées et lianeuses) ou sont épars dans la tige (Monocotylédones typiques).
Cette transformation évolutive part d’une colonne massive qui se creuse d’une moelle, se fragmente et aboutit à des faisceaux individualisés. Elle est en relation avec l’importance de plus en plus grande des feuilles et de leur vascularisation propre. C’est en définitive, la vascularisation foliaire de plus en plus poussée, elle-même corrélative à l’expansion de ces organes, qui justifie cette évolution de la stèle.
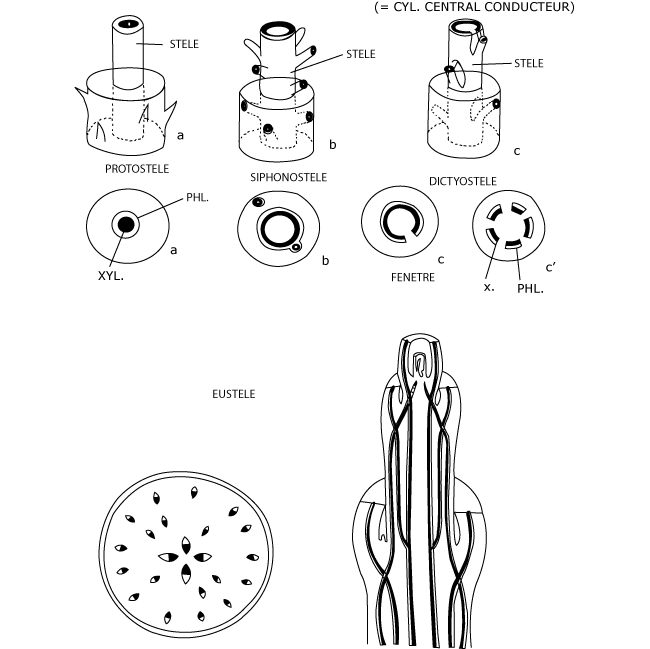 Figure 3 : L’évolution du système vasculaire chez les trachéophytes
Figure 3 : L’évolution du système vasculaire chez les trachéophytes
4. Phylogénèse
Les premiers végétaux vasculaires de la fin du Silurien et de la base du Dévonien se répartissent en trois ensembles distincts les Rhyniophytes (R), les Trimérophytes (T) et les Zostérophyllophytes (Z) réunis en deux lignées évolutives. En effet, les deux premiers constituent la base de l’évolution mégaphylle, le dernier le point de départ des microphylles.
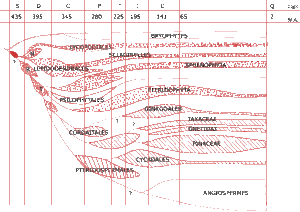 Figure 4 : Origine, importance relative et succession des principaux groupes d’Embryophytes (Bryophytes, Ptéridophytes et Spermatophytes) depuis le Paléozoïque moyen. En pointillé, les ensembles rattachés aux Ptéridophytes s.1; en hacguré, les Gymnospermes; Z = Zostérophyllophytes; R = Rhyniophytes; T = Trimérophytes (M.A = millions d’années)
Figure 4 : Origine, importance relative et succession des principaux groupes d’Embryophytes (Bryophytes, Ptéridophytes et Spermatophytes) depuis le Paléozoïque moyen. En pointillé, les ensembles rattachés aux Ptéridophytes s.1; en hacguré, les Gymnospermes; Z = Zostérophyllophytes; R = Rhyniophytes; T = Trimérophytes (M.A = millions d’années)
Les Trimérophytes possèdent un niveau d’organisation plus avancé que les Rhyniophytes, plus anciennes et plus simples. Les Zostérophyllophytes possèdent des caractères originaux : sporanges latéraux et cordons vasculaires à protoxylème exarche (qui occupe une position périphérique) alors que les deux autres ont des sporanges terminaux et un protoxylème endarche (qui occupe une position centrale). Les Zostérophyllophytes constituent ainsi une lignée très particulière d’où seraient issues toutes les microphylles : Lycopodes, Sélaginelles mais aussi le groupe entièrement fossile des Lépidodendrons comprenant les grands arbres formant les forêts du Dévonien et du Carbonifère.
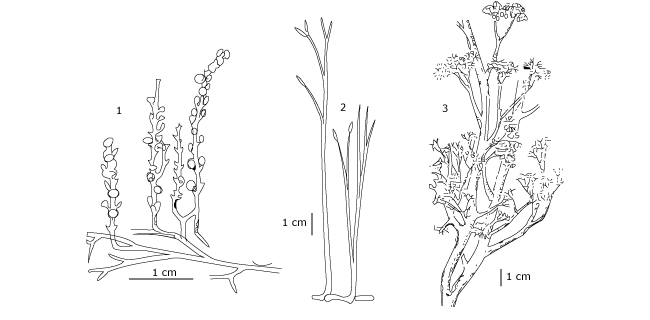 Figure 6 : Reconstitution de fossiles représentatifs des trois divisions des premières plantes vasculaires du Dévonien. 1: Zosterophyllum deciduum (Zostérophyllophyte); 2 : Rhynia major (Rhyniophyte); 3 : Psilophytum dawsonii (Trimérophyte)
Figure 6 : Reconstitution de fossiles représentatifs des trois divisions des premières plantes vasculaires du Dévonien. 1: Zosterophyllum deciduum (Zostérophyllophyte); 2 : Rhynia major (Rhyniophyte); 3 : Psilophytum dawsonii (Trimérophyte)
Malgré la similitude presque complète du mode de reproduction sexuée, il n’y a certainement aucune filiation directe entre Bryophytes et Ptéridophytes. Chez ces dernières, la réduction manifeste du gamétophyte au profit du sporophyte, l’apparition du système complet « racine-tige-feuilles », ainsi que la vascularisation des pousses feuillées sont autant de traits qui éloignent cette lignée de celle des Bryophytes. Aucun argument ne permet de les faire dériver de ces dernières qui, une fois encore, semblent constituer un rameau latéral auquel aucun avenir n’a été réservé.
Il faut encore souligner que la ressemblance physionomique (axes nus à sporanges apicaux) des plus anciens fossiles de Ptéridophytes du Silurien et du Dévonien avec certaines algues ne doit pas faire oublier le fossé profond qui sépare les Chlorophytes les plus avancées des premières plantes vasculaires dont les traces nous sont parvenues.
La lignée des Sphénophytes (y compris les Equisétales) aurait évolué depuis les Rhyniophytes en passant par les Trimérophytes et se distinguerait par une croissance secondaire limitée en raison du fonctionnement des initiales fusiformes. Ce handicap n’a pas empêché les groupes fossiles – tels les Calamites – de produire des arbres imposants à croissance monopodiale munis de vraies feuilles et de racines. Enfin, sont également issues des Trimérophytes, les Psilophytes s.s., les Fougères et toutes les plantes à graines. Les Psilophytes ne constituent à l’heure actuelle qu’un groupe réduit à quelques espèces discrètes (e.a. Psilotum ) alors que les Fougères (Ptéridophytes s.s. ), apparues à la fin du Dévonien, forment encore un groupe fort actuellement de quelque 10.000 espèces occupant tous les habitats et comprenant même des formes arborescentes bien que dépourvues de xylème secondaire. Les botanistes ont longtemps pensé que les Psilophytes, fougère à structure simple avec une tige photosynthétique dépourvue de feuille et poussant depuis un rhizome sans racines, représentaient le groupe le plus primitif dans les Ptéridophytes. Néanmoins, des analyses morphologiques et d’études sur l’ADN ont montré que ce groupe était proche des Ophioglossales (Filicopside) et que sa simplicité de structure pourrait être due à une adaptation à des formes de stress.