Croissance primaire
La croissance de la tige
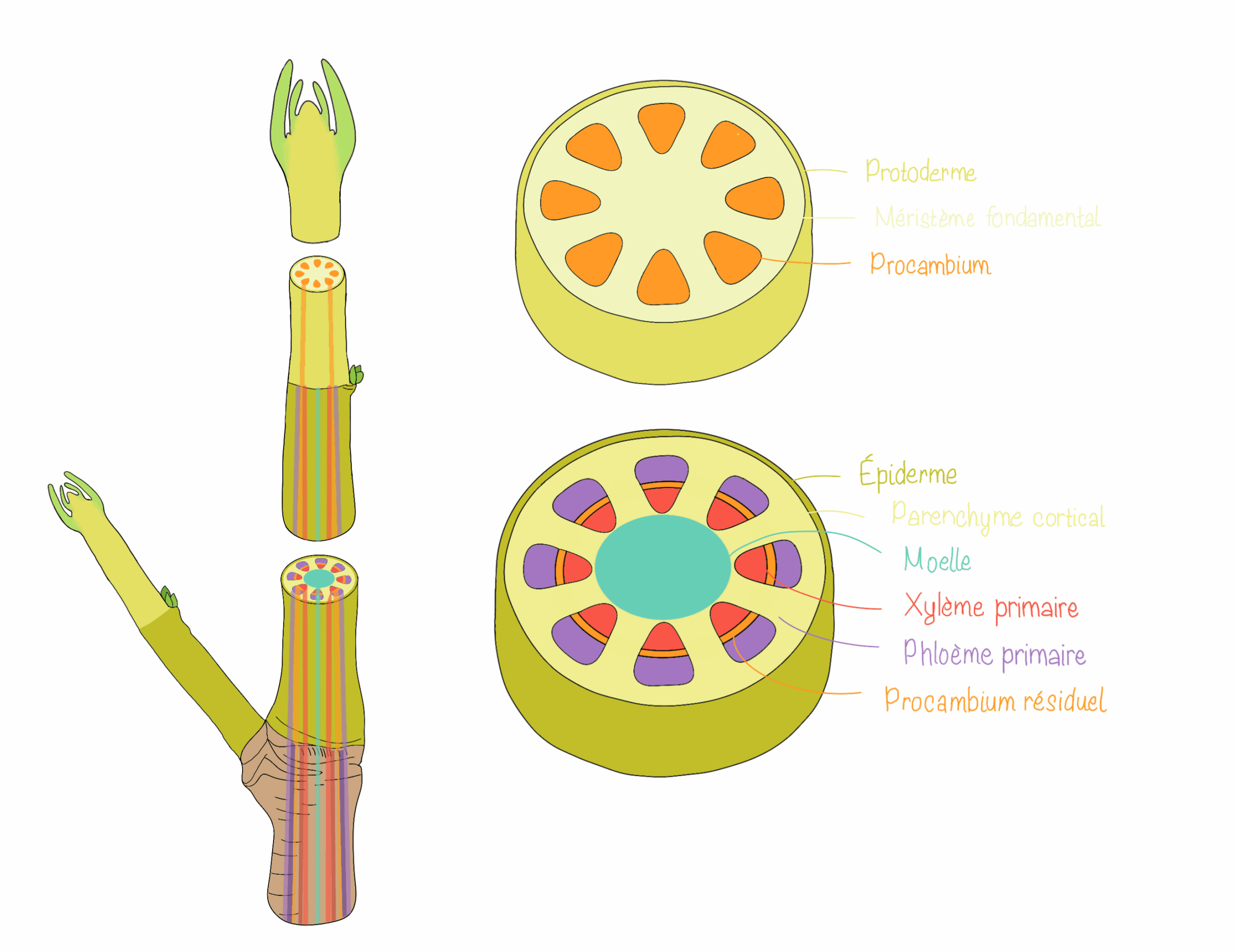
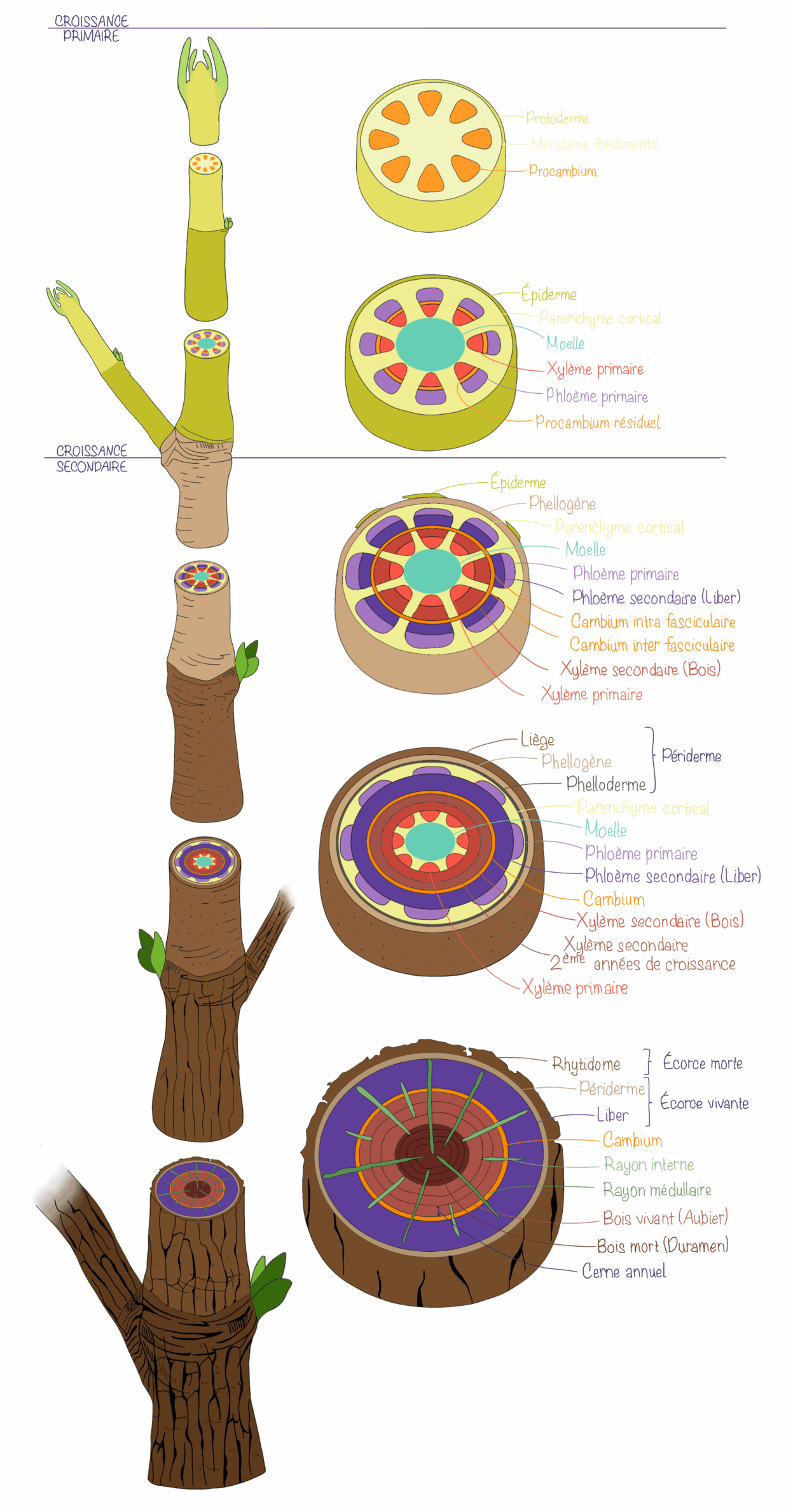
La tige croît grâce à l’activité du méristème apical caulinaire, qui produit des cellules se différenciant ensuite en différents tissus primaires. Comme dans la racine, le protoderme se différencie en épiderme qui protège la tige. Sous l’épiderme, le méristème fondamental donne naissance au collenchyme, au parenchyme cortical et à la moelle centrale, des tissus de soutien, remplissage et stockage. Le procambium forme les tissus conducteurs primaires, c’est-à-dire le xylème primaire et le phloème primaire qui s’organisent selon l’architecture propre à chaque type de plante (monocotylée vs. dicotylée). Cette croissance primaire établit l’organisation interne de la tige, essentielle à la conduction de la sève et au maintien de sa structure mécanique.
Structure interne de la tige chez les dicotylées
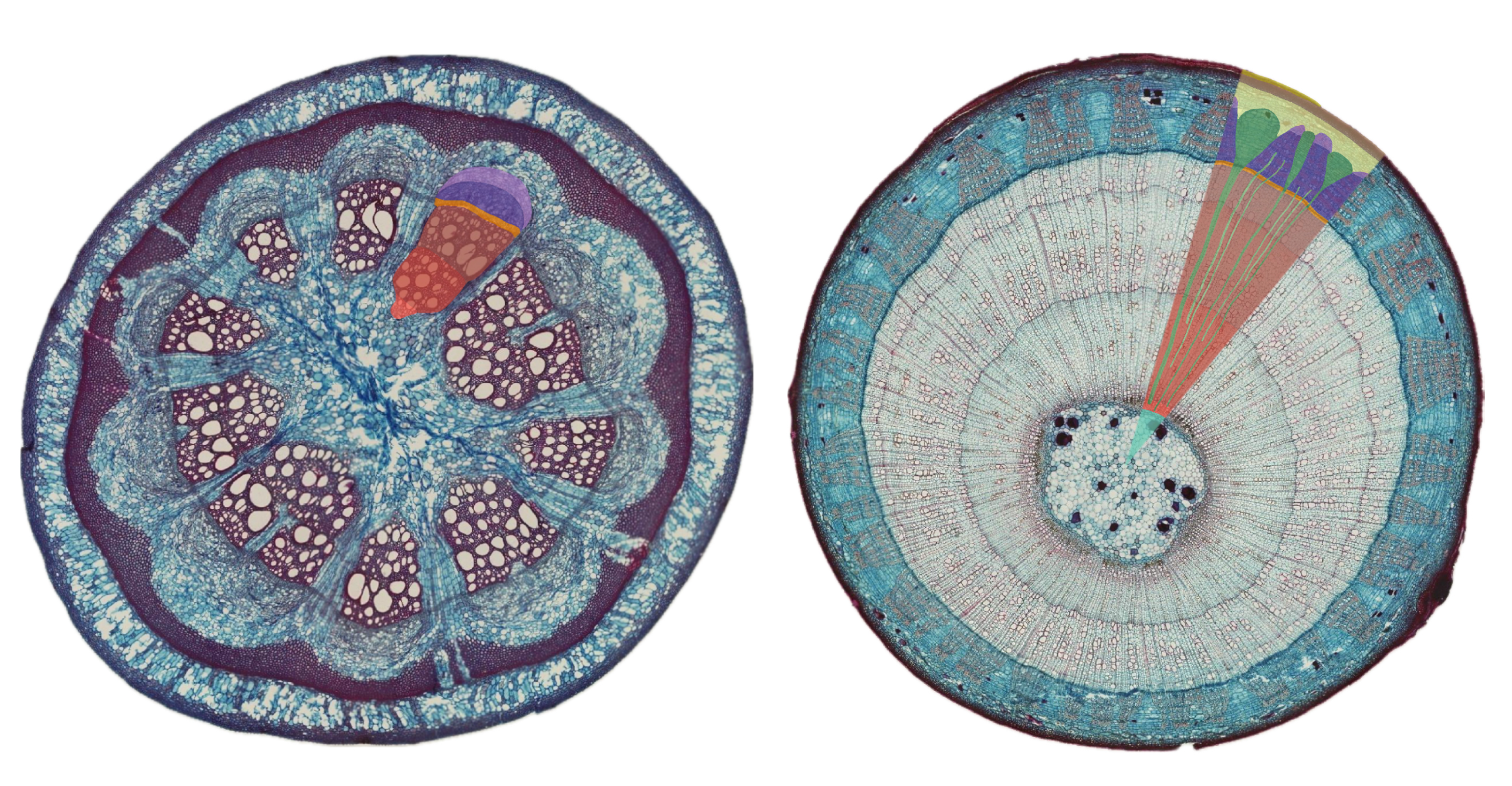
Chez les tiges de dicotylées, la structure interne peut se présenter sous deux formes principales.
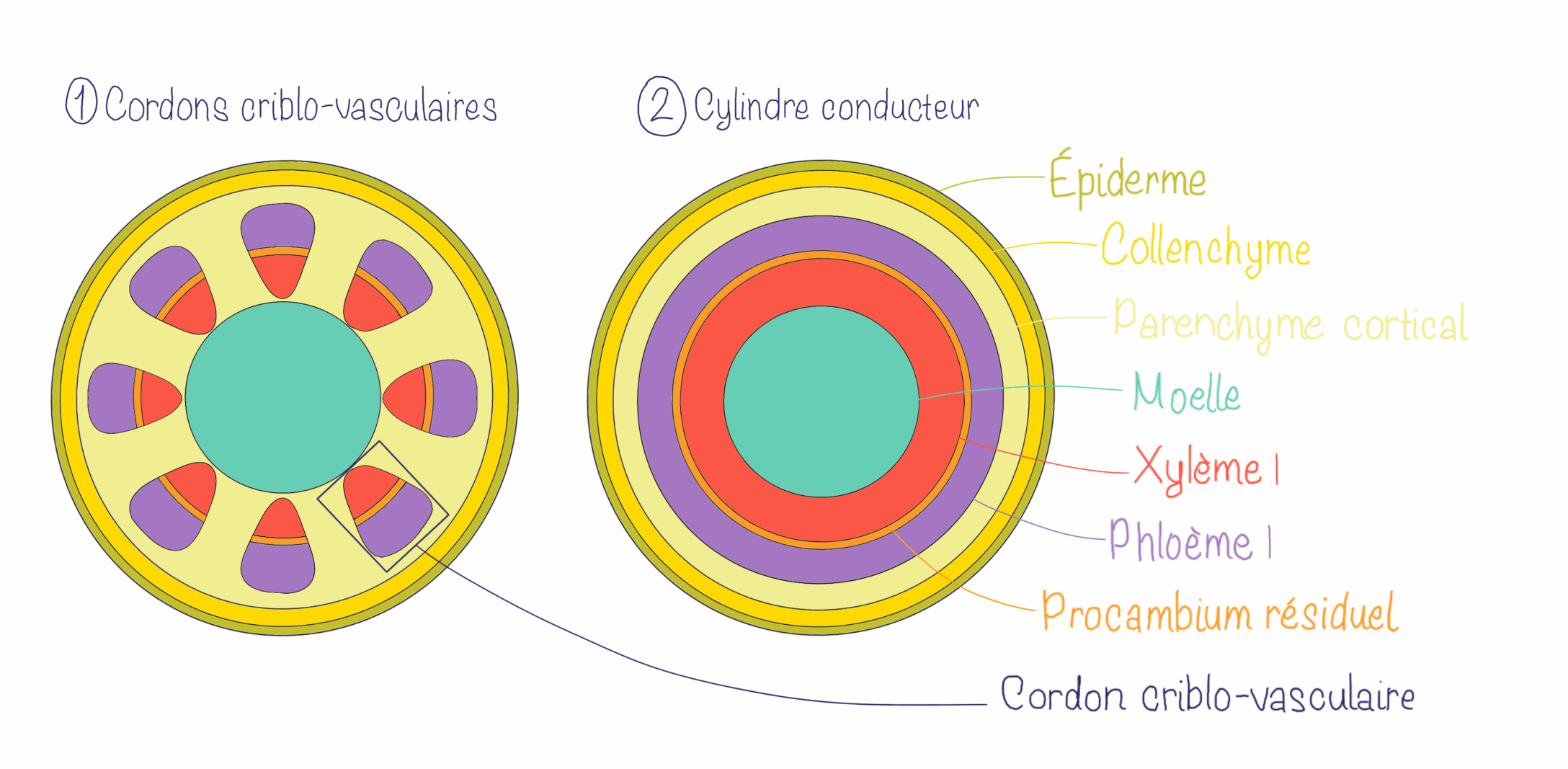
Dans l’organisation en cylindre conducteur, les tissus conducteurs sont regroupés en anneaux autour d’une moelle centrale, le xylème se trouvant vers l’intérieur et le phloème vers l’extérieur. Il y a donc une séparation claire au niveau du parenchyme en parenchyme cortical (cortex) et médullaire (moelle).
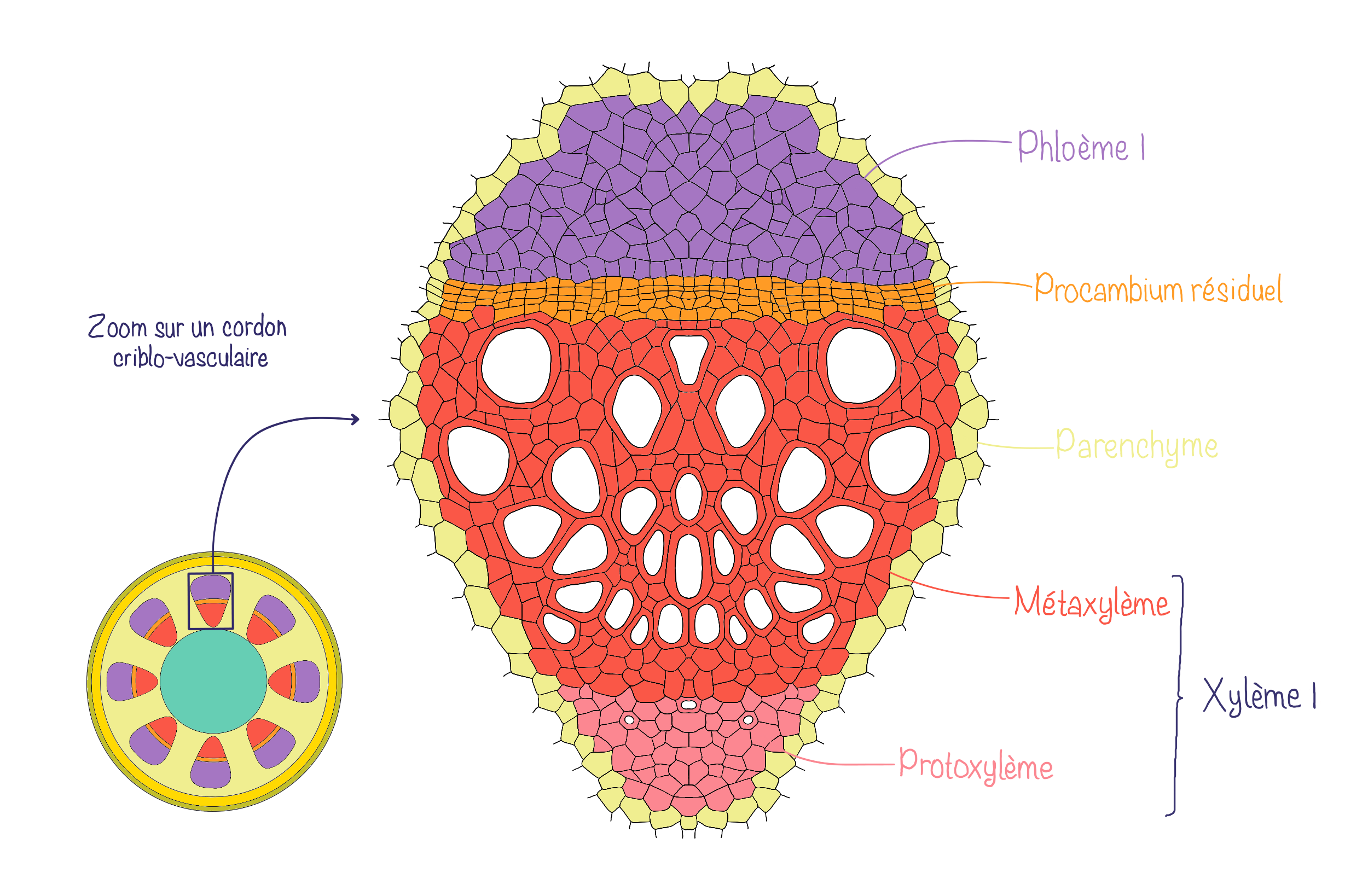
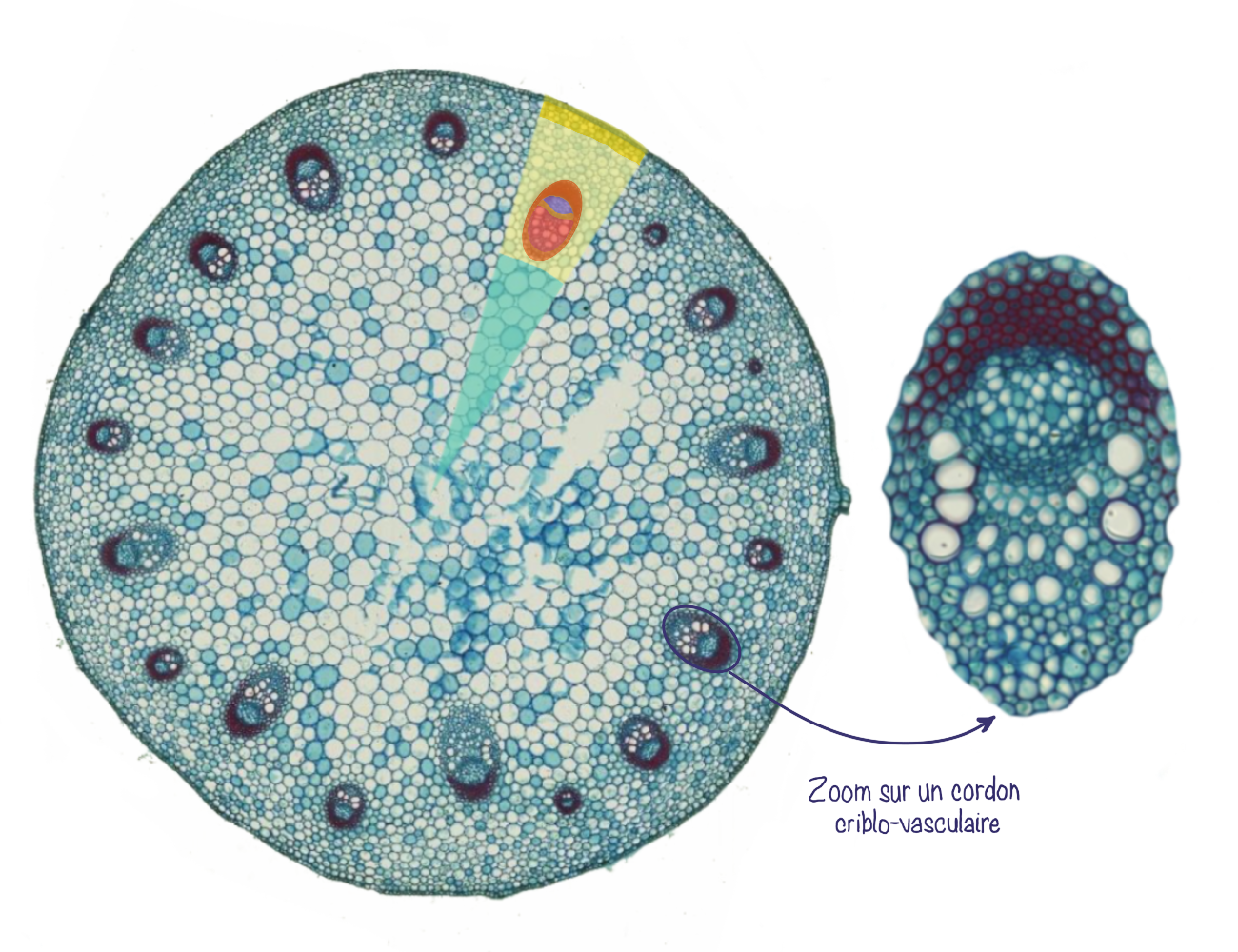
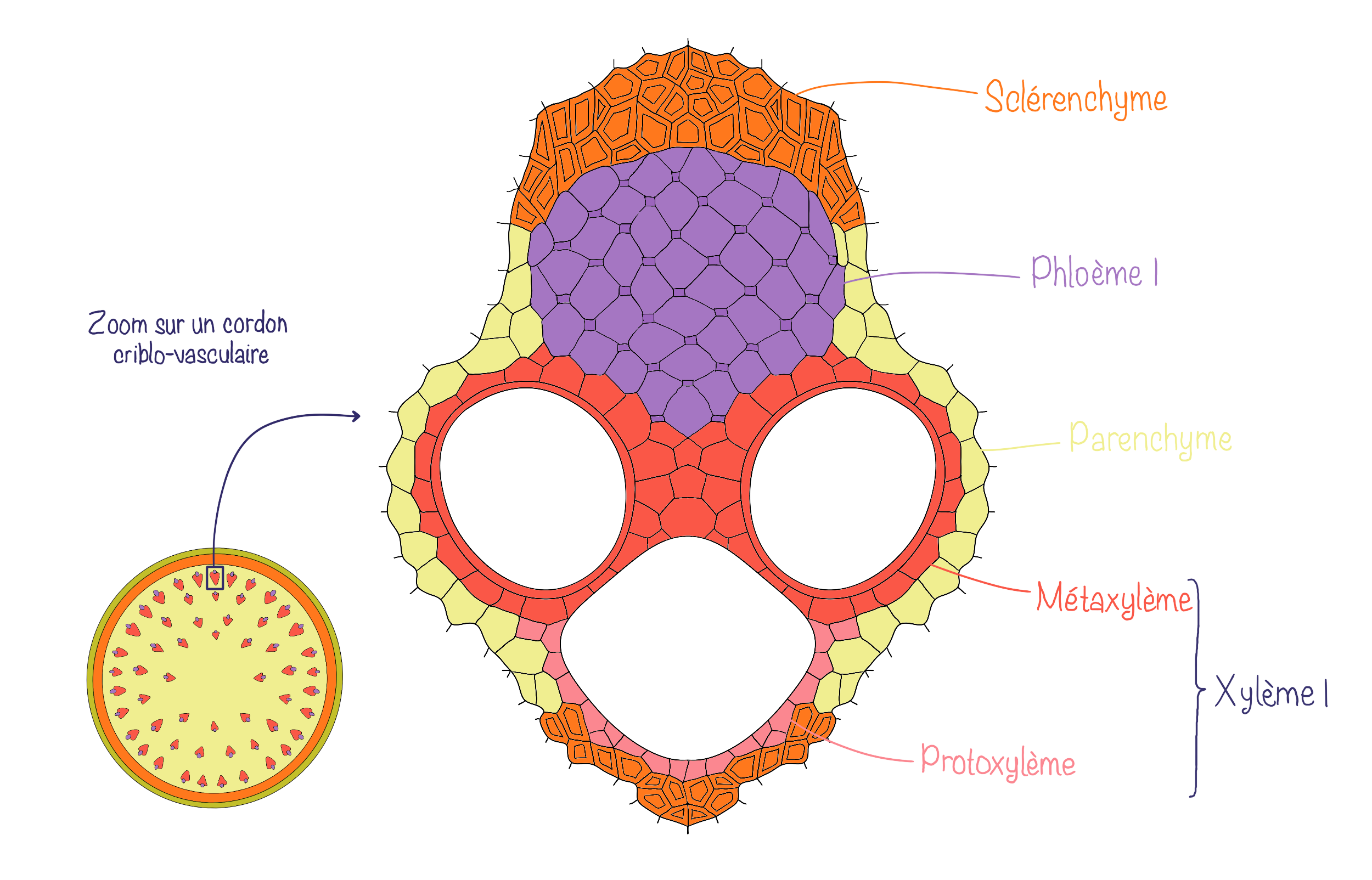
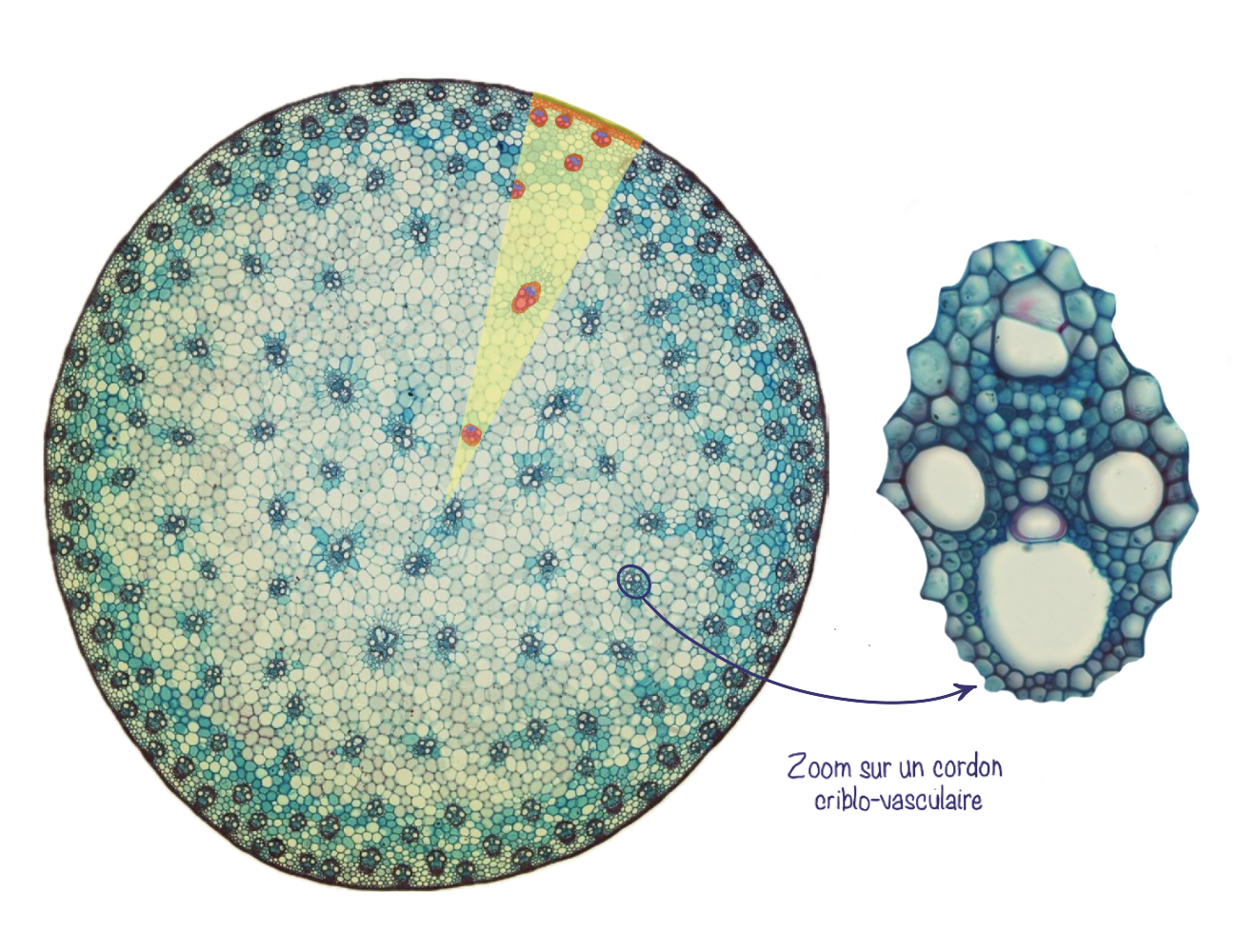
Dans l’organisation en faisceaux conducteurs ou cordons criblo-vasculaires, les tissus conducteurs forment plusieurs faisceaux conducteurs (ou cordons criblo-vasculaires) qui sont séparés par du parenchyme inter-fasciculaire (aussi appelé rayons parenchymateux). Le parenchyme est toujours divisé en parenchyme cortical (cortex) et médullaire (moelle) mais il est continu via les rayons. Au sein des cordons criblo-vasculaires, la position relative du xylème et phloème est la même que pour le cylindre conducteur (xylème vers l’intérieur et phloème vers l’extérieur).
Dans les 2 cas, le xylème et le phloème sont séparés par du procambium résiduel.
Structure interne de la tige chez les monocotylées
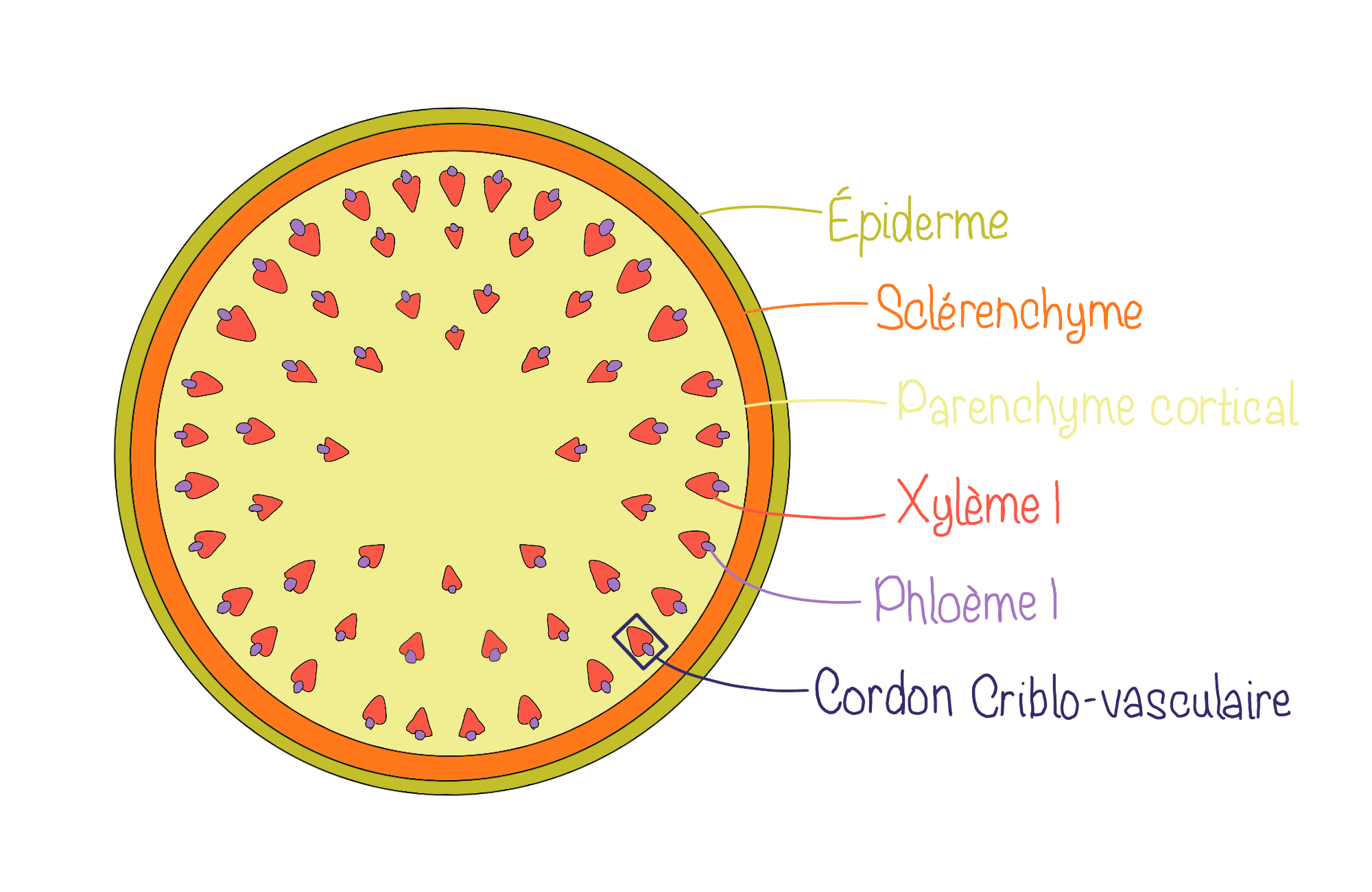
L’organisation interne des tiges de monocotylées se caractérise par la présence de cordons criblo-vasculaires dispersés de manière irrégulière dans le parenchyme fondamental. Contrairement aux tiges de dicotylées, il n’y a pas de moelle centrale bien délimitée. De plus, ces faisceaux ne possèdent pas de procambium résiduel. Enfin, sous l’épiderme, le collenchyme, typique des dicotylées, est remplacé par du sclérenchyme, qui assure un rôle de soutien plus rigide.
Croissance secondaire
Evolution de la croissance I vers la croissance II de la tige
Comme dans la racine, la croissance secondaire de la tige permet son épaississement et son renforcement mécanique. Le cambium se met en place à partir des cellules du procambium résiduel, qui se différencient et se divisent jusqu’à former un manchon circulaire. Dans l’organisation en cordons criblo-vasculaires (ou faisceaux conducteurs), ce cercle se constitue par la jonction du cambium intrafasciculaire, situé dans les cordons, et du cambium interfasciculaire, issu des zones entre les cordons. Dans l’organisation en cylindre continu, en revanche, le procambium forme déjà un cylindre cambial homogène.
Ce cambium produit ensuite du bois (xylème secondaire) vers l’intérieur et du liber (phloème secondaire) vers l’extérieur, assurant à la fois conduction et soutien.
Parallèlement, le phellogène se développe à partir de certaines cellules du parenchyme proches de l’épiderme. Cette assise de cellules engendre du phelloderme vers l’intérieur et du liège vers l’extérieur, qui constitue le périderme. Cette nouvelle couche protectrice remplace progressivement l’épiderme, qui se dégrade et disparaît au cours du développement.
Structure de la tige secondaire
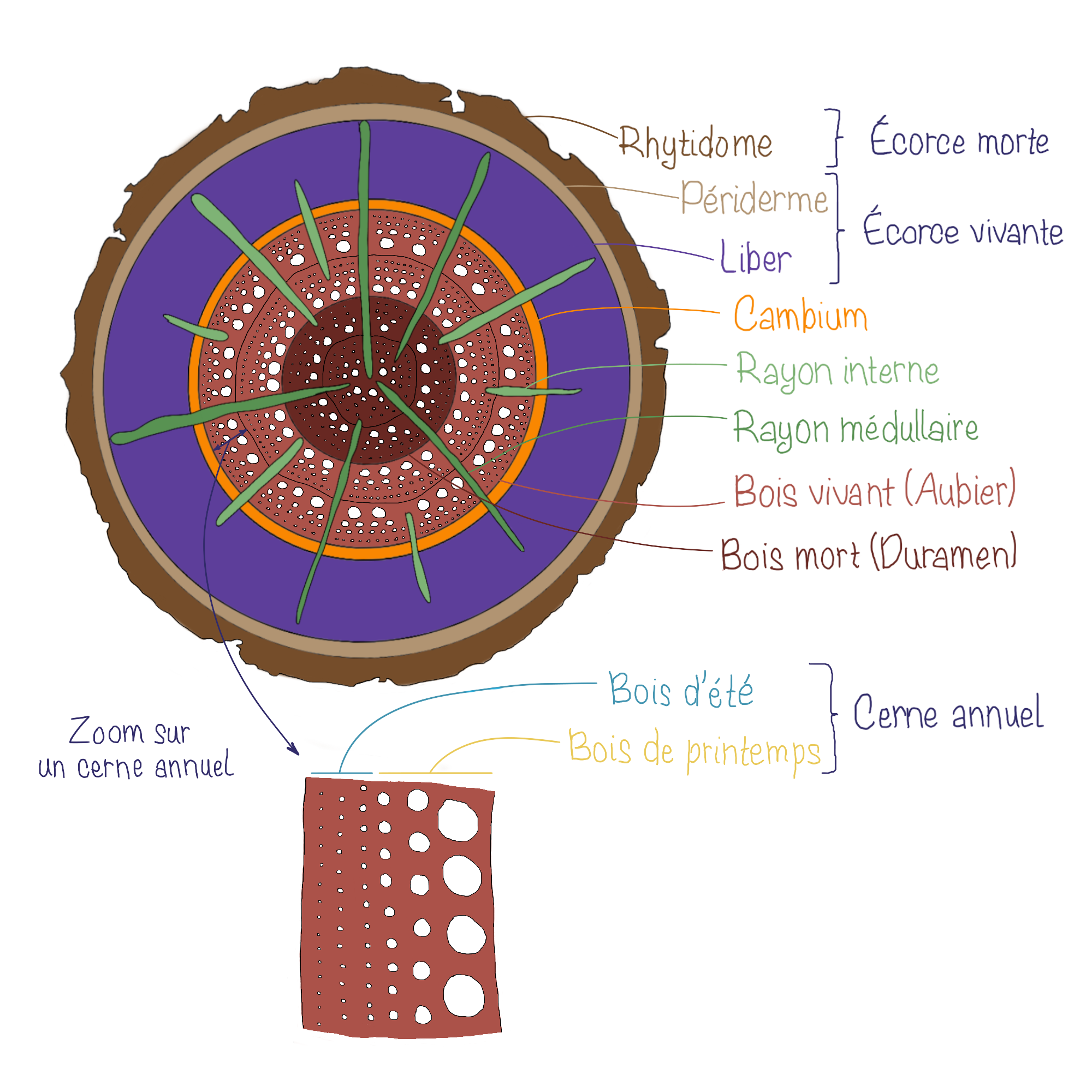
Le périderme et le rhytidome
Le périderme est perforé par des lenticelles, des petites structures spécialisées permettant les échanges gazeux entre l’air et les tissus vivants sous-jacents.
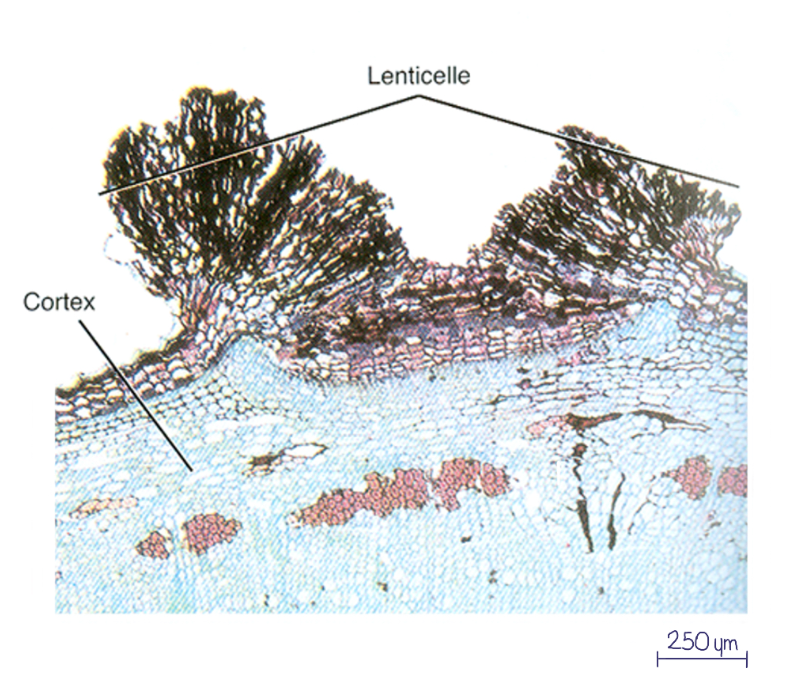
Avec les années, de nouvelles assises de phellogène se forment plus en profondeur. Les couches externes meurent et s’accumulent pour former le rhytidome, surface externe visible à la surface des arbres adultes. Le rhytidome est formé par la superposition de péridermes successifs. L’écorce quant à elle regroupe tous les tissus extérieurs au cambium (liber, phloème I, cortex, périderme(s)).
Le bois
L’activité cambiale est influencée par les saisons : au printemps, le xylème secondaire formé est composé de larges vaisseaux (bois de printemps), dont la conductivité pour la sève est importante, tandis qu’en été, il devient plus dense et épaissit (éléments de vaisseaux plus étroits, plus grande proportion de fibres, bois d’été), renforçant le soutien mécanique. Cette alternance entre bois de printemps et d’été donne naissance aux cernes annuels, visibles en coupe transversale et permettant d’estimer l’âge d’un arbre.
Avec le temps, l’accumulation continue de couches de xylème secondaire repousse le liber vers l’extérieur et comprime progressivement les tissus primaires, qui finissent par disparaître. L’essentiel de la masse de la tige adulte est alors constitué de bois. Cependant seule la zone périphérique de celui-ci reste physiologiquement active : il s’agit de l’aubier, qui assure la conduction de la sève brute et conserve encore des cellules vivantes. Plus au centre, les couches anciennes de xylème secondaire perdent leur fonction conductrice et forment le duramen. Ce bois, plus sombre et plus dense, est inerte mais assure la rigidité et la durabilité de la tige.
Certaines cellules cambiales se différencient également en cellule de rayons. Ces structures radiales composées de tissu parenchymateux traversent le bois et l’écorce pour assurer la conduction horizontale de l’eau, des nutriments et des réserves. On distingue les rayons médullaires, qui s’étendent sur toute la longueur du tronc et relient le centre aux couches périphériques, et les rayons internes, plus courts, qui ne sont pas connectés directement au centre.