1. Les méristèmes
Au cours de l’embryogenèse chez les plantes, une structure simplifiée est mise en place, qui comprend deux populations de ‘cellules-souches’ – le méristème caulinaire et le méristème racinaire. Ces cellules-souches indifférenciés appelés méristèmes se multiplient activement.
Toutes les structures post-embryonnaires des végétaux dérivent de l’activité de ces méristèmes, qui sont donc des groupes de cellules peu spécialisées, peu différencies, qui se divisent et dont les cellules dérivées élaborent les organes et les tissus. Grâce aux méristèmes, la plante possède en permanence un état embryonnaire (tout au moins jusqu’à la transition florale pour le méristème caulinaire) ce qui justifie l’étude de son développement au travers de son ontogenèse (histoire du développement de l’individu à partir du germe).
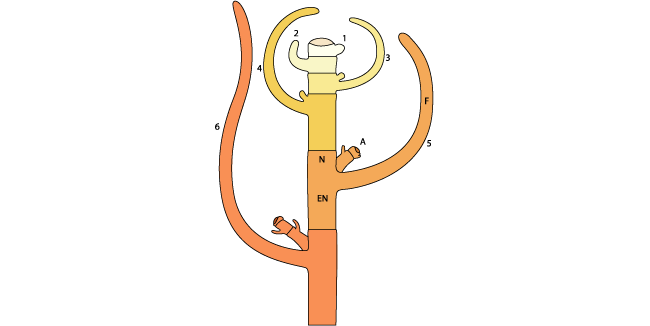
A la fin de l’embryogenèse, la plantule possède l’organisation de base de la plante : méristème racinaire et méristème caulinaire en continuité, bauches foliaires. L’appareil arien de la plante se développe par addition de nouvelles unités, selon un mode itratif. Au niveau de la tige, chaque unit est constituée d’un nœud comprenant une feuille et son bourgeon axillaire, ainsi que l’entre-nœud correspondant (Figure 1). Parmi les différentes espèces végétales, une grande variabilité de formes existe sur base de ce schéma. Ce développement itératif permet une importante amplification du nombre de méristèmes disponibles et la possibilités de reconstituer tout ou partie de portions endommages, faculté particulièrement importante pour un organisme fixé, qui ne peut guère s’échapper face aux contraintes environnementales biotiques et abiotiques.
B. Les méristèmes primaires
On reconnaît deux principaux types de méristèmes primaires : le méristème racinaire et le méristème caulinaire. Deux différences essentielles caractérisent ces deux types de méristèmes :- selon le rôle :
- selon la différenciation :
- le méristème caulinaire de l’embryon met en place directement des méristèmes caulinaires axillaires (qui font partie du développement programmé de la plante) ; dans ce cas, il n’y a pas de dédifférenciation. Il existe également des méristèmes adventifs qui peuvent apparaître secondairement mais pas à un endroit habituel. Ils se forment alors par dédifférenciation de cellules différencies (exemple des racines adventives apparaissant lors du bouturage).
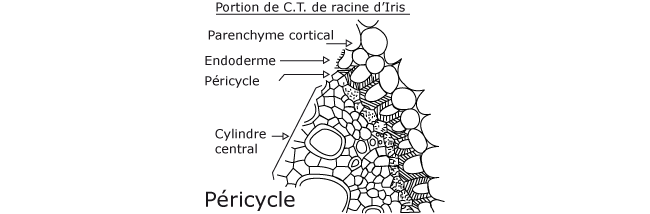
- concernant les racines, les ramifications apparaissent lorsque la différenciation est djà termine. Pour donner une racine secondaire, il se forme donc obligatoirement un nouveau méristème par dédifférenciation des cellules, généralement provenant du péricycle (Figure 2).
2. Le méristème racinaire
Le méristème apical de racine est lui aussi formé durant l’embryogenèse. Il élabore les tissus de la racine et la coiffe: il est uniquement histogène. Il ne produit pas d’organes latéraux et n’est donc pas organogène. Les racines latérales se forment de manière endogène à quelque distance de l’apex. Leur méristème se différencie à partir du péricycle (assise cellulaire la plus externe, située entre l’écorce et la stèle ou partie centrale conductrice de la racine). Le péricycle (Figure 2) initiée les ramifications de la racine. La structure et le fonctionnement des ramifications sont identiques à celui du méristème apical de la racine. L’ensemble des méristèmes racinaires édifie les organes souterrains de la plante constituant le système racinaire. N.B.: Il existe des exceptions car il y a des racines aériennes et il y a des tiges souterraines.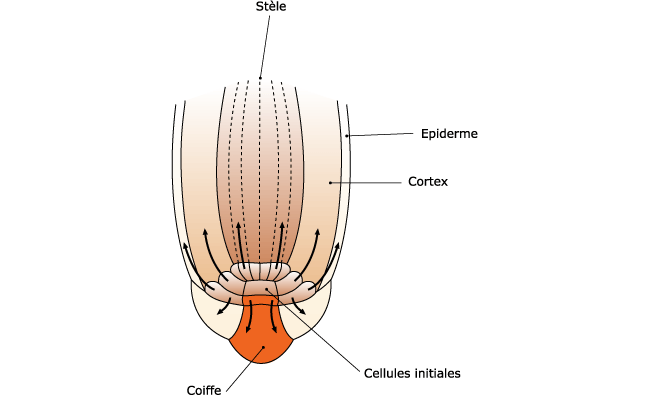

A. Structure
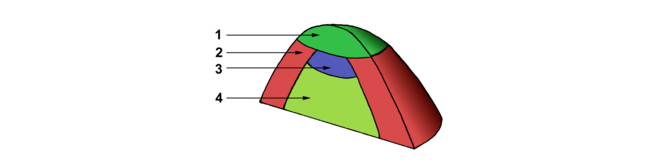
Bien que de formes et de tailles très différentes, les méristèmes caulinaires présentent peu de diversité quant à leur structure interne. Leur organisation peut être décrite comme étant la superposition de 2 patrons: l’un en couches cellulaires; l’autre en zones. 1) Le méristème caulinaire apical est stratifié en couches cellulaires superposées reconnues dès 1868 par Hanstein. La couche épidermique L1, constituée d’une unique assise cellulaire, recouvre des couches cellulaires internes distinctes L2 et L3 qui peuvent chacune comprendre plus d’une assise cellulaire (Figure 5).
- la tunica comprenant une ou plusieurs assises superficielles dans lesquelles les cellules se divisent quasi exclusivement dans le sens anticline (perpendiculaire à la surface).
- le corpus sous-jacent qui est constitué de cellules se divisant dans toutes les directions.
- la zone centrale (ZC) comprend des cellules assez vacuolisées, pauvres en ARN et protéines, dont les noyaux ont de petits nucléoles. Ces cellules se divisent relativement rarement.
- la zone périphérique (ZP), de forme plus ou moins annulaire, entoure la ZC. Les cellules y sont de taille plus petite qu’en ZC. Elles sont plus riches en ARN et protéines. Le cytoplasme y est dense et les vacuoles très petites. Les noyaux occupent presque tout le volume cellulaire et les nucléoles sont de grande taille. Cette zone est le siège d’une activité mitotique importante et de synthèses actives d’ADN.
- le méristème médullaire (MM), situé sous les deux autres zones, est constitué de cellules qui, en section longitudinale, apparaissent aplaties. Elles sont très vacuolisées et contiennent des grains d’amidon relativement gros. Elles se divisent presque exclusivement de façon péricline. L’activité mitotique y est plus réduite. Cette zone, qui ressemble à un cambium, est plus active et plus développée chez les espèces caulescentes que chez les espèces en rosette.